

An endogenous aryl hydrocarbon receptor ligand, ITE, induces regulatory T cells and ameliorates experimental colitis
- PMID: 29672155
- PMCID: PMC6139639
- DOI: 10.1152/ajpgi.00413.2017
An endogenous aryl hydrocarbon receptor ligand, ITE, induces regulatory T cells and ameliorates experimental colitis
Abstract
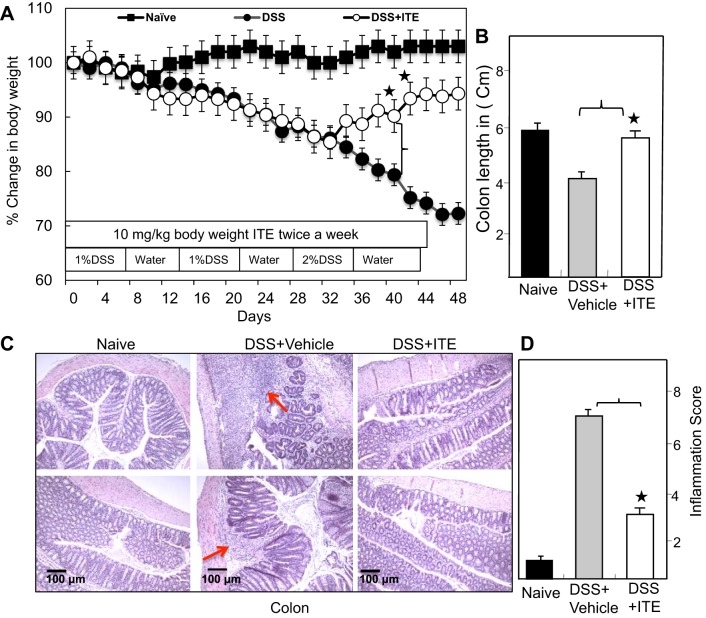
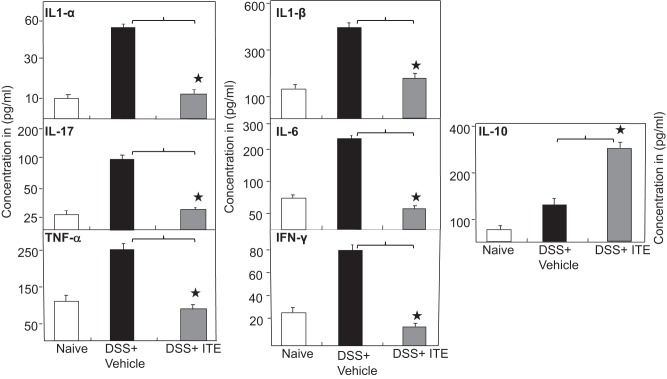
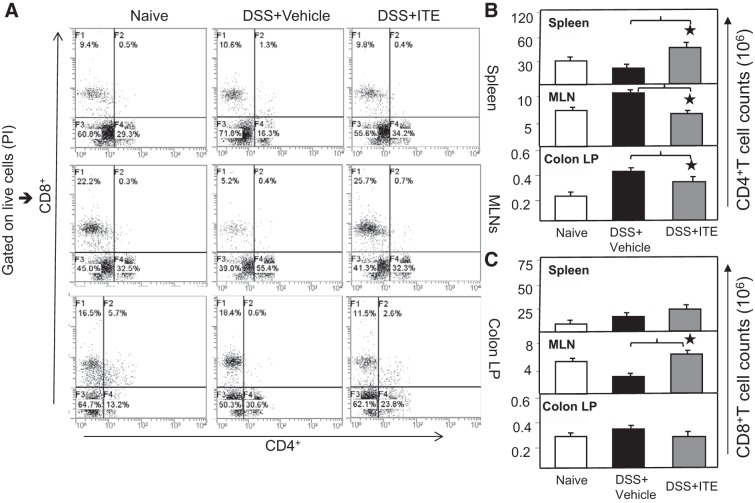
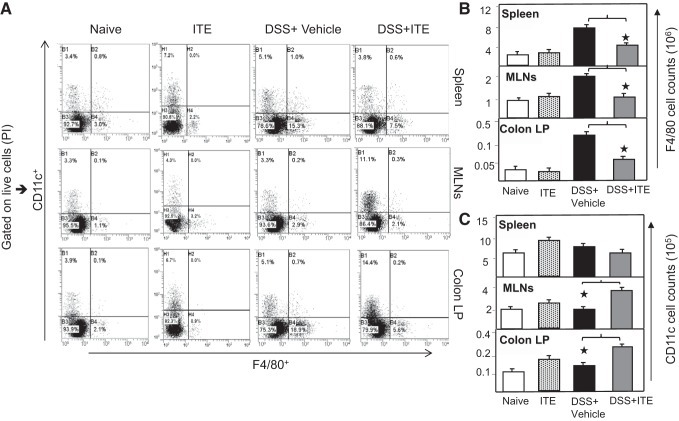
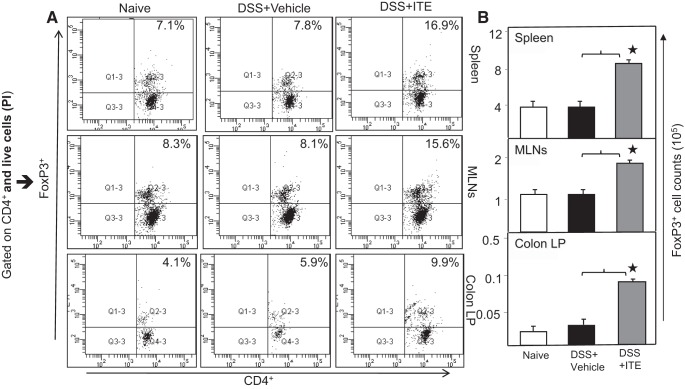
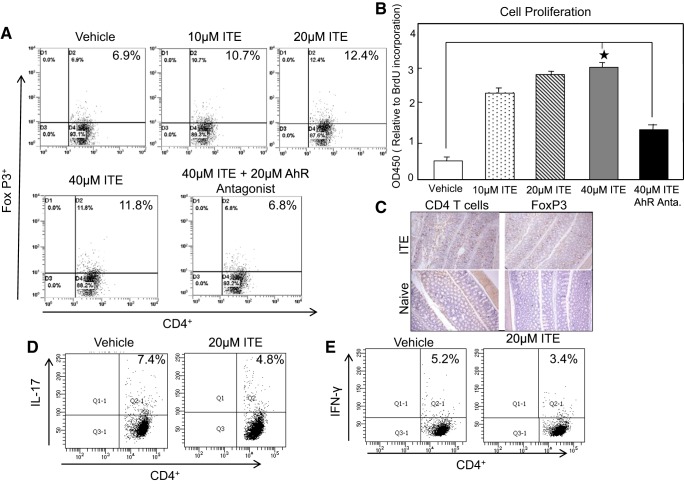
Inflammatory bowel disease (IBD) is a chronic intestinal inflammatory condition that affects millions of people with high morbidity and health care costs. The precise etiology of IBD is unknown, but clear evidence suggests that intestinal inflammation is caused by an excessive immune response to mucosal antigens. Recent studies have shown that activation of the aryl hydrocarbon receptor (AhR) induces regulatory T cells (Tregs) and suppresses autoimmune diseases. In the current study, we investigated if a nontoxic ligand of AhR, 2-(1' H-indole-3'-carbonyl)-thiazole-4-carboxylic acid methyl ester (ITE), can attenuate dextran sodium sulfate-induced colitis. Our studies demonstrated that in mice that received ITE treatment in vivo, colitis pathogenesis, including a decrease in body weight, was significantly reversed along with the systemic and intestinal inflammatory cytokines. ITE increased the expression of Tregs in spleen, mesenteric lymph nodes (MLNs), and colon lamina propria lymphocytes (cLPL) of mice with colitis when compared with controls. This induction of Tregs was reversed by AhR antagonist treatment in vitro. ITE treatment also increased dendritic cells (CD11c+) and decreased macrophages (F4/80+) from the spleen, MLNs, and cLPL in mice with colitis. ITE also reversed the systemic and intestinal frequency of CD4+ T cells during colitis and suppressed inflammatory cytokines including IFN-γ, TNF-α, IL-17, IL-6, and IL-1 as well as induced IL-10 levels. These findings suggest that ITE attenuates colitis through induction of Tregs and reduction in inflammatory CD4+ T cells and cytokines. Therefore, our work demonstrates that the nontoxic endogenous AhR ligand ITE may serve as a therapeutic modality to treat IBD. NEW & NOTEWORTHY We report the novel finding that activation of the aryl hydrocarbon receptor with the nontoxic ligand 2-(1'H-indole-3'-carbonyl)-thiazole-4-carboxylic acid methyl ester (ITE) induces regulatory T cells (Tregs) and suppresses inflammatory bowel disease (IBD). Our data suggest that ITE diminishes colitis pathology through induction of Tregs; reduces inflammatory cytokines, inflammation score, and macrophage frequency; and induces DCs resulting in amelioration of colitis. Therefore, nontoxic endogenous ITE promotes the induction of Tregs and may be useful for the treatment of IBD.
Keywords: Crohn’s disease; Th1/Th17; inflammatory bowel disease; ulcerative colitis.
Figures
References
-
- Bacchetta R, Passerini L, Gambineri E, Dai M, Allan SE, Perroni L, Dagna-Bricarelli F, Sartirana C, Matthes-Martin S, Lawitschka A, Azzari C, Ziegler SF, Levings MK, Roncarolo MG. Defective regulatory and effector T cell functions in patients with FOXP3 mutations. J Clin Invest 116: 1713–1722, 2006. doi: 10.1172/JCI25112. - DOI - PMC - PubMed
-
- Barrett JC, Hansoul S, Nicolae DL, Cho JH, Duerr RH, Rioux JD, Brant SR, Silverberg MS, Taylor KD, Barmada MM, Bitton A, Dassopoulos T, Datta LW, Green T, Griffiths AM, Kistner EO, Murtha MT, Regueiro MD, Rotter JI, Schumm LP, Steinhart AH, Targan SR, Xavier RJ, Libioulle C, Sandor C, Lathrop M, Belaiche J, Dewit O, Gut I, Heath S, Laukens D, Mni M, Rutgeerts P, Van Gossum A, Zelenika D, Franchimont D, Hugot JP, de Vos M, Vermeire S, Louis E, Cardon LR, Anderson CA, Drummond H, Nimmo E, Ahmad T, Prescott NJ, Onnie CM, Fisher SA, Marchini J, Ghori J, Bumpstead S, Gwilliam R, Tremelling M, Deloukas P, Mansfield J, Jewell D, Satsangi J, Mathew CG, Parkes M, Georges M, Daly MJ; NIDDK IBD Genetics Consortium; Belgian-French IBD Consortium; Wellcome Trust Case Control Consortium . Genome-wide association defines more than 30 distinct susceptibility loci for Crohn’s disease. Nat Genet 40: 955–962, 2008. doi: 10.1038/ng.175. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials