

ROS and Trypanosoma cruzi: Fuel to infection, poison to the heart
- PMID: 29672619
- PMCID: PMC5908069
- DOI: 10.1371/journal.ppat.1006928
ROS and Trypanosoma cruzi: Fuel to infection, poison to the heart
Abstract
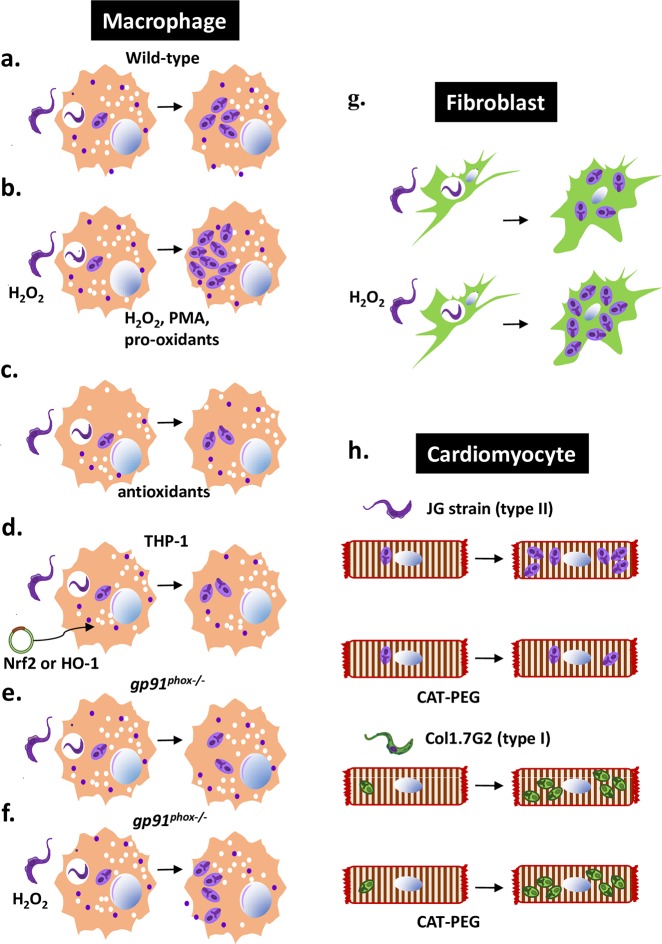
The activation of macrophage respiratory burst in response to infection with Trypanosoma cruzi inflicts oxidative damage to the host's tissues. For decades, the role of reactive oxygen species (ROS) in the elimination of T. cruzi was taken for granted, but recent evidence suggests parasite growth is stimulated in oxidative environments. It is still a matter of debate whether indeed oxidative environments provide ideal conditions (e.g., iron availability in macrophages) for T. cruzi growth and whether indeed ROS signals directly to stimulate growth. Nitric oxide (NO) and ROS combine to form peroxynitrite, participating in the killing of phagocytosed parasites by activated macrophages. In response to infection, mitochondrial ROS are produced by cardiomyocytes. They contribute to oxidative damage that persists at the chronic stage of infection and is involved in functional impairment of the heart. In this review, we discuss how oxidative stress helps parasite growth during the acute stage and how it participates in the development of cardiomyopathy at the chronic stage.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
-
- Baehner RL, Karnovsky ML. Deficiency of reduced nicotinamide-adenine dinucleotide oxidase in chronic granulomatous disease. Science. 1968;162(3859):1277–9. . - PubMed
-
- Murray HW. Pretreatment with phorbol myristate acetate inhibits macrophage activity against intracellular protozoa. J Reticuloendothel Soc. 1982;31(6):479–87. . - PubMed
-
- Locksley RM, Klebanoff SJ. Oxygen-dependent microbicidal systems of phagocytes and host defense against intracellular protozoa. J Cell Biochem. 1983;22(3):173–85. doi: 10.1002/jcb.240220306 . - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
