

Pathogen-derived extracellular vesicles mediate virulence in the fatal human pathogen Cryptococcus gattii
- PMID: 29674675
- PMCID: PMC5908794
- DOI: 10.1038/s41467-018-03991-6
Pathogen-derived extracellular vesicles mediate virulence in the fatal human pathogen Cryptococcus gattii
Abstract
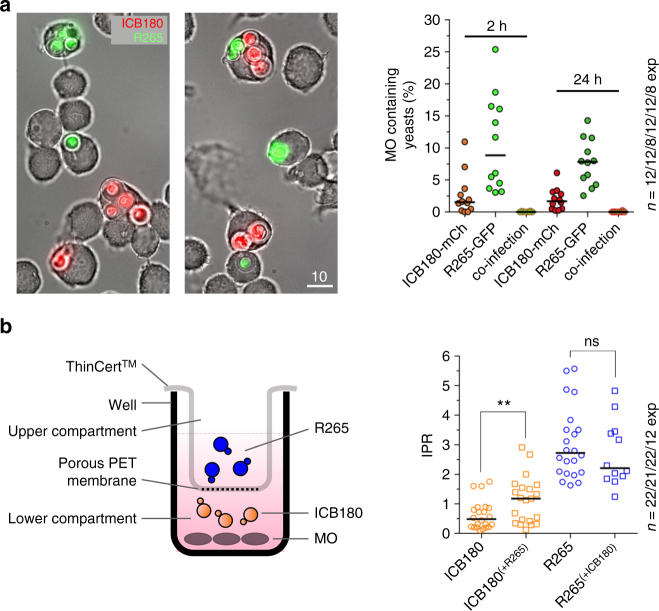
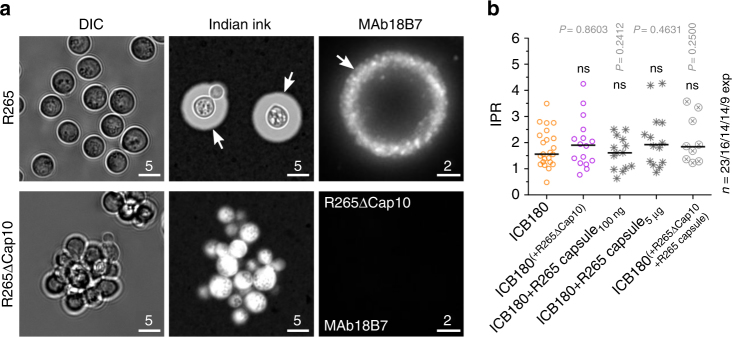
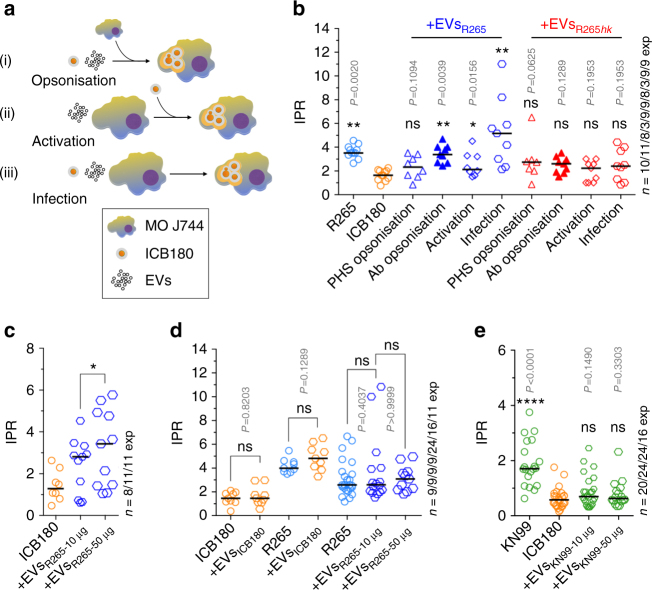
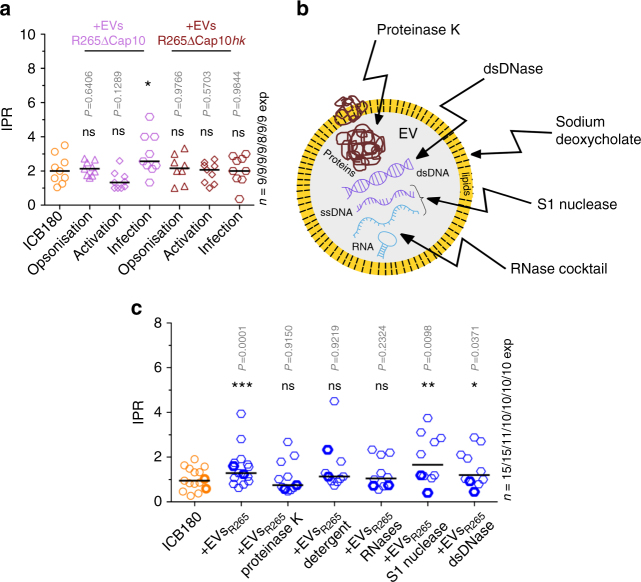
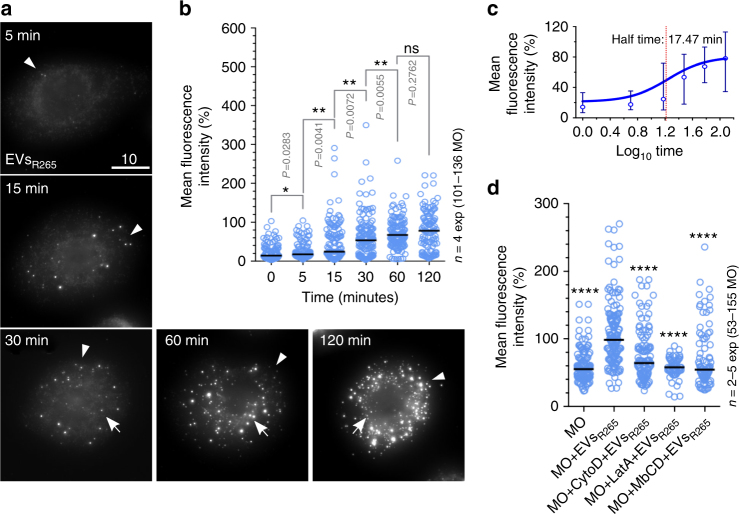
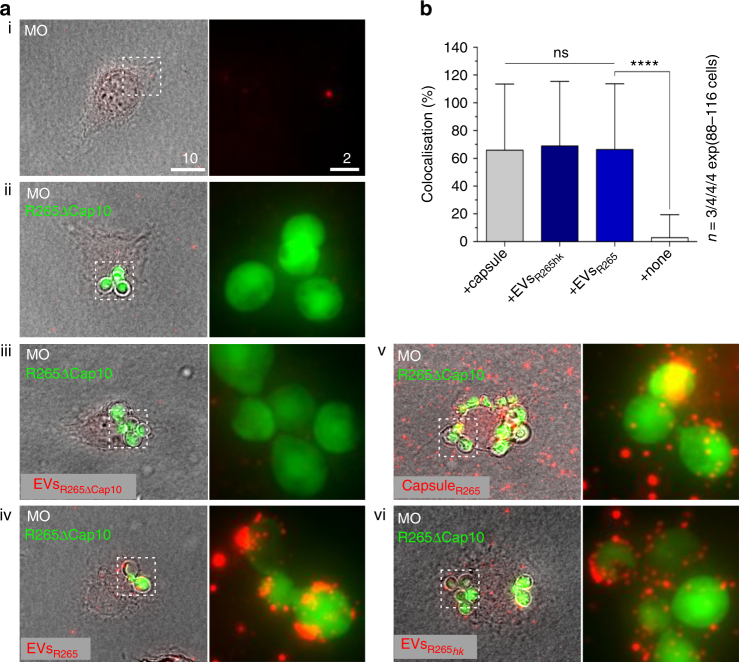
The Pacific Northwest outbreak of cryptococcosis, caused by a near-clonal lineage of the fungal pathogen Cryptococcus gattii, represents the most significant cluster of life-threatening fungal infections in otherwise healthy human hosts currently known. The outbreak lineage has a remarkable ability to grow rapidly within human white blood cells, using a unique 'division of labour' mechanism within the pathogen population, where some cells adopt a dormant behaviour to support the growth of neighbouring cells. Here we demonstrate that pathogenic 'division of labour' can be triggered over large cellular distances and is mediated through the release of extracellular vesicles by the fungus. Isolated vesicles released by virulent strains are taken up by infected host macrophages and trafficked to the phagosome, where they trigger the rapid intracellular growth of non-outbreak fungal cells that would otherwise be eliminated by the host. Thus, long distance pathogen-to-pathogen communication via extracellular vesicles represents a novel mechanism to control complex virulence phenotypes in Cryptococcus gattii and, potentially, other infectious species.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
