

Exosomal circRNA_100284 from arsenite-transformed cells, via microRNA-217 regulation of EZH2, is involved in the malignant transformation of human hepatic cells by accelerating the cell cycle and promoting cell proliferation
- PMID: 29674685
- PMCID: PMC5908808
- DOI: 10.1038/s41419-018-0485-1
Exosomal circRNA_100284 from arsenite-transformed cells, via microRNA-217 regulation of EZH2, is involved in the malignant transformation of human hepatic cells by accelerating the cell cycle and promoting cell proliferation
Abstract
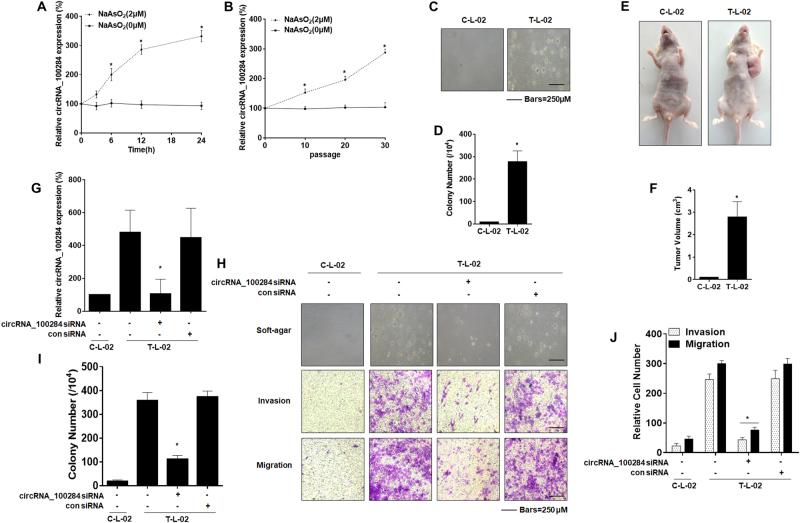
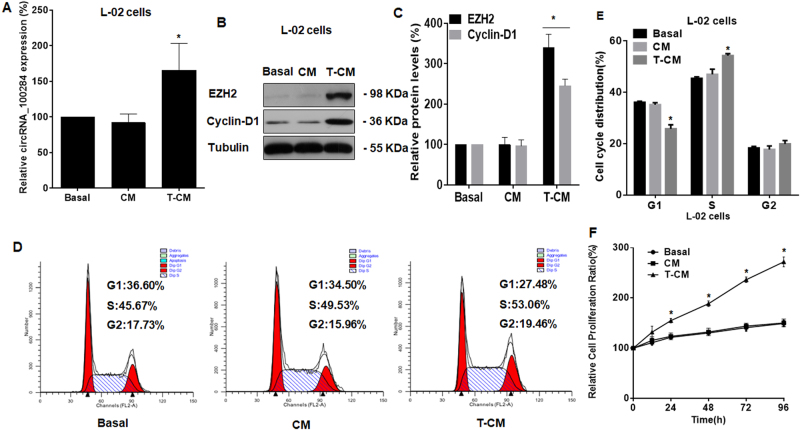
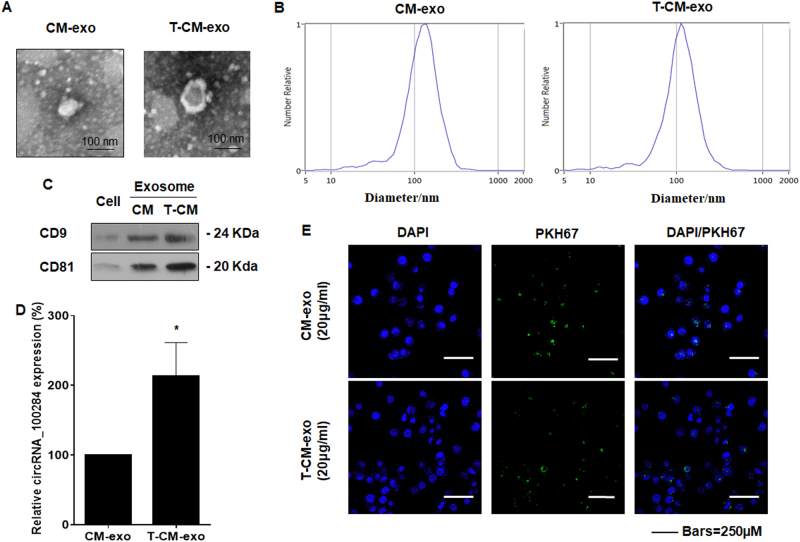
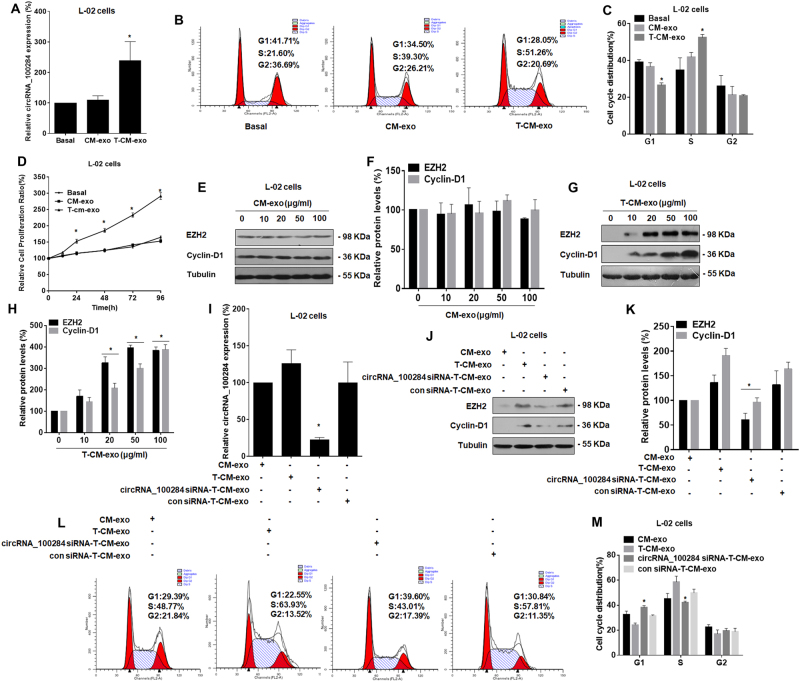
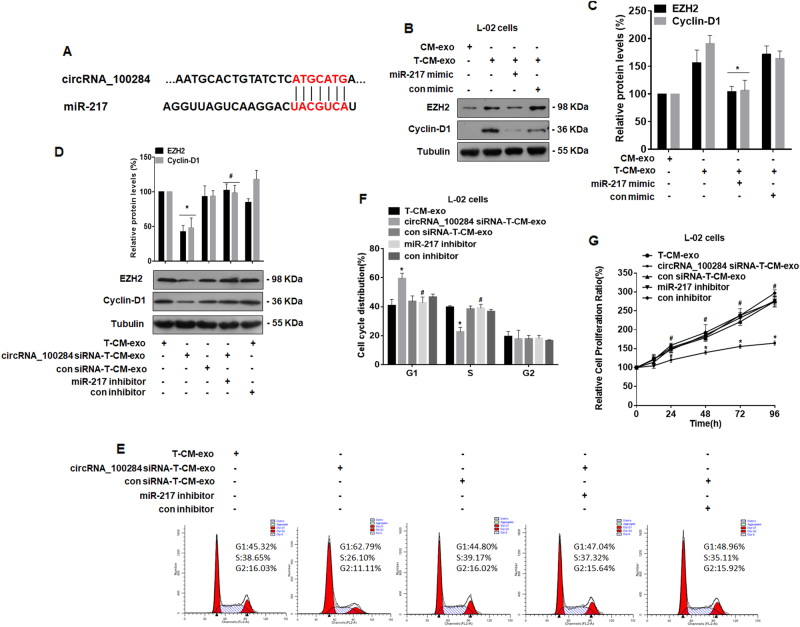
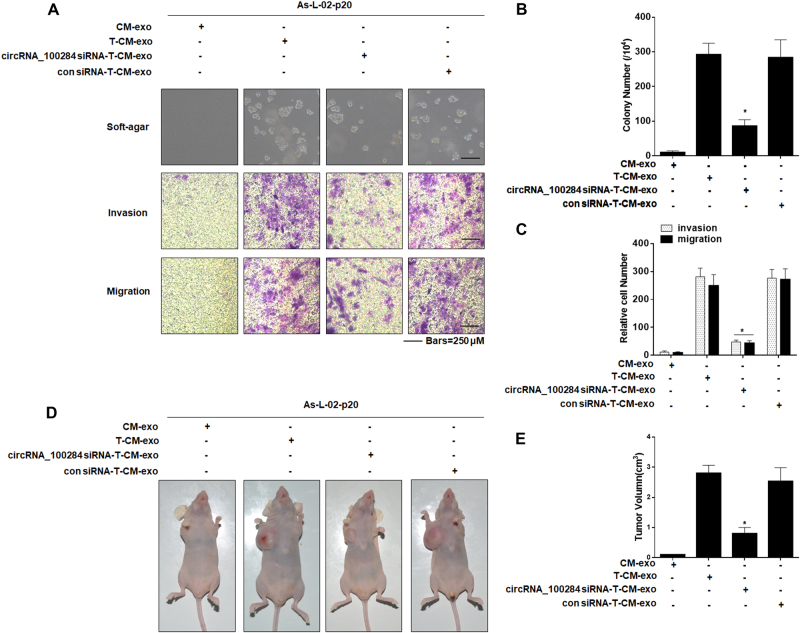
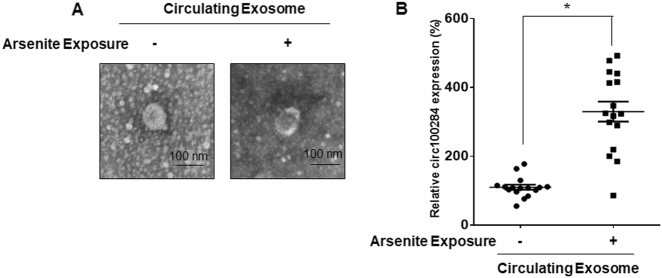
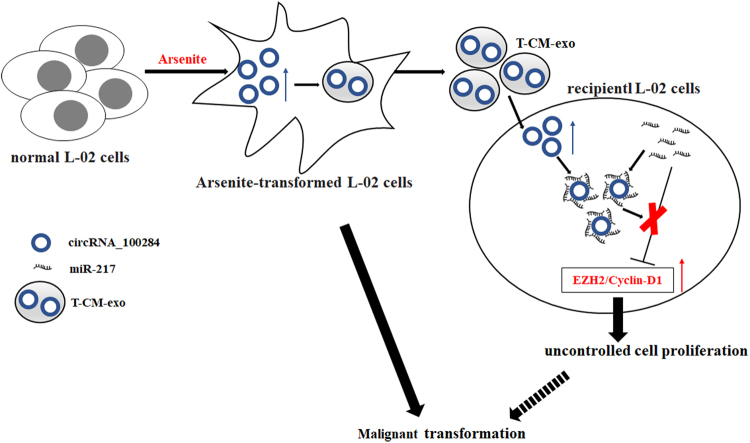
Intercellular communication between malignant cells and neighboring nonmalignant cells is involved in carcinogenesis. In the progression of carcinogenesis, exosomes are messengers for intercellular communication. Circular RNAs (circRNAs) are noncoding RNAs with functions that include regulation of the cell cycle and proliferation. However, the functions of exosomal circRNAs are not clear. The present research aimed to determine whether circRNAs secreted from arsenite-transformed human hepatic epithelial (L-02) cells are transferred into normal L-02 cells and become functionally active in the normal cells. The results showed that circRNA_100284 is involved in the malignant transformation of L-02 cells induced by arsenite. The medium from transformed L-02 cells induced upregulation of circRNA_100284, accelerated the cell cycle, and promoted proliferation of normal L-02 cells. Transformed cells transferred circRNA_100284 into normal L-02 cells via exosomes and led to the malignant transformation of the non-transformed cells. Knockdown of circRNA_100284, which reduced circRNA_100284 levels in exosomes derived from transformed L-02 cells, blocked the accelerated cell cycle and reduced proliferation and malignancy. In addition, in normal L-02 cells, exosomal circRNA_100284 derived from arsenite-transformed L-02 cells induced acceleration of the cell cycle and promoted proliferation via acting as a sponge of microRNA-217. Further, exosomal circRNA_100284 was upregulated in the sera of people exposed to arsenite. Thus, exosomes derived from transformed L-02 cells transferred circRNA_100284 to surrounding cells, which induced an accelerated cell cycle and promoted proliferation of normal liver cells and led to the malignant transformation of the non-transformed cells. The findings support the concept that exosomal circRNAs are involved in cell-cell communication during carcinogenesis induced by arsenite.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
