

Enzymes Involved in Posttranscriptional RNA Metabolism in Gram-Negative Bacteria
- PMID: 29676246
- PMCID: PMC5912700
- DOI: 10.1128/microbiolspec.RWR-0011-2017
Enzymes Involved in Posttranscriptional RNA Metabolism in Gram-Negative Bacteria
Abstract
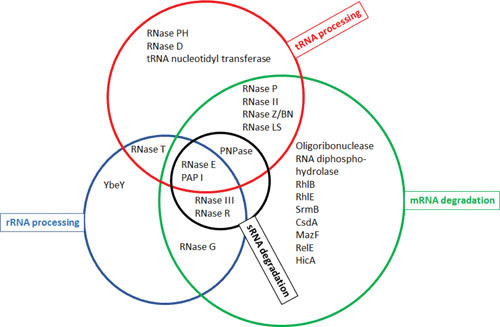
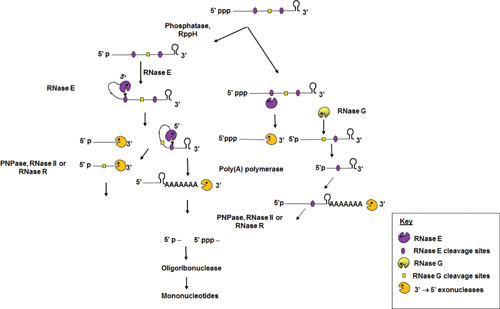
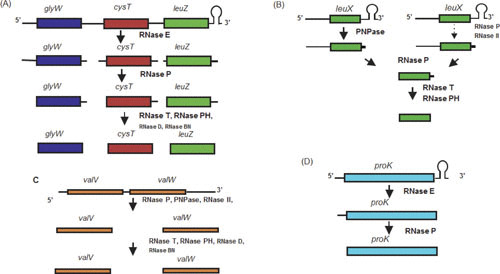
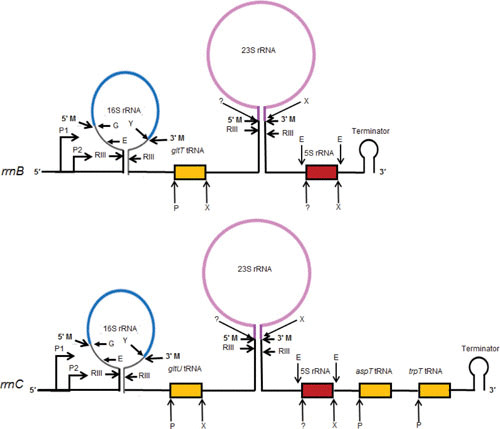
Gene expression in Gram-negative bacteria is regulated at many levels, including transcription initiation, RNA processing, RNA/RNA interactions, mRNA decay, and translational controls involving enzymes that alter translational efficiency. In this review, we discuss the various enzymes that control transcription, translation, and RNA stability through RNA processing and degradation. RNA processing is essential to generate functional RNAs, while degradation helps control the steady-state level of each individual transcript. For example, all the pre-tRNAs are transcribed with extra nucleotides at both their 5' and 3' termini, which are subsequently processed to produce mature tRNAs that can be aminoacylated. Similarly, rRNAs that are transcribed as part of a 30S polycistronic transcript are matured to individual 16S, 23S, and 5S rRNAs. Decay of mRNAs plays a key role in gene regulation through controlling the steady-state level of each transcript, which is essential for maintaining appropriate protein levels. In addition, degradation of both translated and nontranslated RNAs recycles nucleotides to facilitate new RNA synthesis. To carry out all these reactions, Gram-negative bacteria employ a large number of endonucleases, exonucleases, RNA helicases, and poly(A) polymerase, as well as proteins that regulate the catalytic activity of particular RNases. Under certain stress conditions, an additional group of specialized endonucleases facilitate the cell's ability to adapt and survive. Many of the enzymes, such as RNase E, RNase III, polynucleotide phosphorylase, RNase R, and poly(A) polymerase I, participate in multiple RNA processing and decay pathways.
Figures
Similar articles
-
The critical role of RNA processing and degradation in the control of gene expression.FEMS Microbiol Rev. 2010 Sep;34(5):883-923. doi: 10.1111/j.1574-6976.2010.00242.x. Epub 2010 Jun 24. FEMS Microbiol Rev. 2010. PMID: 20659169 Review.
-
The Phosphorolytic Exoribonucleases Polynucleotide Phosphorylase and RNase PH Stabilize sRNAs and Facilitate Regulation of Their mRNA Targets.J Bacteriol. 2016 Nov 18;198(24):3309-3317. doi: 10.1128/JB.00624-16. Print 2016 Dec 15. J Bacteriol. 2016. PMID: 27698082 Free PMC article.
-
RNA components of Escherichia coli degradosome: evidence for rRNA decay.Proc Natl Acad Sci U S A. 1998 Mar 17;95(6):3157-61. doi: 10.1073/pnas.95.6.3157. Proc Natl Acad Sci U S A. 1998. PMID: 9501232 Free PMC article.
-
The C-terminal half of RNase E, which organizes the Escherichia coli degradosome, participates in mRNA degradation but not rRNA processing in vivo.Mol Microbiol. 1999 Jul;33(1):188-99. doi: 10.1046/j.1365-2958.1999.01465.x. Mol Microbiol. 1999. PMID: 10411735
-
The Escherichia coli RNA degradosome: structure, function and relationship in other ribonucleolytic multienzyme complexes.Biochem Soc Trans. 2002 Apr;30(2):150-5. Biochem Soc Trans. 2002. PMID: 12035760 Review.
Cited by
-
Genes Differentially Expressed by Haemophilus ducreyi during Anaerobic Growth Significantly Overlap Those Differentially Expressed during Experimental Infection of Human Volunteers.J Bacteriol. 2022 May 17;204(5):e0000522. doi: 10.1128/jb.00005-22. Epub 2022 Apr 4. J Bacteriol. 2022. PMID: 35377183 Free PMC article.
-
Endonucleolytic processing plays a critical role in the maturation of ribosomal RNA in Methanococcus maripaludis.RNA Biol. 2023 Jan;20(1):760-773. doi: 10.1080/15476286.2023.2258035. Epub 2023 Sep 20. RNA Biol. 2023. PMID: 37731260 Free PMC article.
-
Neisseria meningitidis Sibling Small Regulatory RNAs Connect Metabolism with Colonization by Controlling Propionate Use.J Bacteriol. 2023 Mar 21;205(3):e0046222. doi: 10.1128/jb.00462-22. Epub 2023 Mar 1. J Bacteriol. 2023. PMID: 36856428 Free PMC article.
-
RNase III and RNase E Influence Posttranscriptional Regulatory Networks Involved in Virulence Factor Production, Metabolism, and Regulatory RNA Processing in Bordetella pertussis.mSphere. 2021 Aug 25;6(4):e0065021. doi: 10.1128/mSphere.00650-21. Epub 2021 Aug 18. mSphere. 2021. PMID: 34406853 Free PMC article.
-
Prevalence of small base-pairing RNAs derived from diverse genomic loci.Biochim Biophys Acta Gene Regul Mech. 2020 Jul;1863(7):194524. doi: 10.1016/j.bbagrm.2020.194524. Epub 2020 Mar 5. Biochim Biophys Acta Gene Regul Mech. 2020. PMID: 32147527 Free PMC article. Review.
References
-
- Ono M, Kuwano M. 1979. A conditional lethal mutation in an Escherichia coli strain with a longer chemical lifetime of messenger RNA. J Mol Biol 129:343–357. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
