

Nuclear Import Receptor Inhibits Phase Separation of FUS through Binding to Multiple Sites
- PMID: 29677513
- PMCID: PMC6234985
- DOI: 10.1016/j.cell.2018.03.003
Nuclear Import Receptor Inhibits Phase Separation of FUS through Binding to Multiple Sites
Abstract
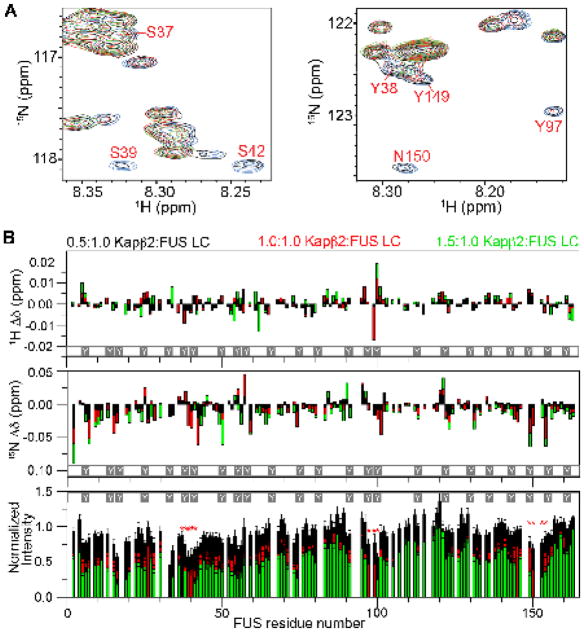
Liquid-liquid phase separation (LLPS) is believed to underlie formation of biomolecular condensates, cellular compartments that concentrate macromolecules without surrounding membranes. Physical mechanisms that control condensate formation/dissolution are poorly understood. The RNA-binding protein fused in sarcoma (FUS) undergoes LLPS in vitro and associates with condensates in cells. We show that the importin karyopherin-β2/transportin-1 inhibits LLPS of FUS. This activity depends on tight binding of karyopherin-β2 to the C-terminal proline-tyrosine nuclear localization signal (PY-NLS) of FUS. Nuclear magnetic resonance (NMR) analyses reveal weak interactions of karyopherin-β2 with sequence elements and structural domains distributed throughout the entirety of FUS. Biochemical analyses demonstrate that most of these same regions also contribute to LLPS of FUS. The data lead to a model where high-affinity binding of karyopherin-β2 to the FUS PY-NLS tethers the proteins together, allowing multiple, distributed weak intermolecular contacts to disrupt FUS self-association, blocking LLPS. Karyopherin-β2 may act analogously to control condensates in diverse cellular contexts.
Keywords: FUS; PY-NLS; RNA granule; amyotrophic lateral sclerosis; biomolecular condensate; intrinsically disordered protein; karyopherin-β2; liquid-liquid phase separation; low-complexity sequences; transportin-1.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
Phase Separation of FUS Is Suppressed by Its Nuclear Import Receptor and Arginine Methylation.Cell. 2018 Apr 19;173(3):706-719.e13. doi: 10.1016/j.cell.2018.03.004. Cell. 2018. PMID: 29677514
-
Mechanism of karyopherin-β2 binding and nuclear import of ALS variants FUS(P525L) and FUS(R495X).Sci Rep. 2021 Feb 12;11(1):3754. doi: 10.1038/s41598-021-83196-y. Sci Rep. 2021. PMID: 33580145 Free PMC article.
-
Structural and energetic basis of ALS-causing mutations in the atypical proline-tyrosine nuclear localization signal of the Fused in Sarcoma protein (FUS).Proc Natl Acad Sci U S A. 2012 Jul 24;109(30):12017-21. doi: 10.1073/pnas.1207247109. Epub 2012 Jul 9. Proc Natl Acad Sci U S A. 2012. PMID: 22778397 Free PMC article.
-
Effect of nuclear import receptors on liquid-liquid phase separation.Biophys Physicobiol. 2020 Feb 20;17:25-29. doi: 10.2142/biophysico.BSJ-2019052. eCollection 2020. Biophys Physicobiol. 2020. PMID: 33110735 Free PMC article. Review.
-
Transportin-1 and Transportin-2: protein nuclear import and beyond.FEBS Lett. 2014 May 21;588(10):1857-68. doi: 10.1016/j.febslet.2014.04.023. Epub 2014 Apr 26. FEBS Lett. 2014. PMID: 24780099 Review.
Cited by
-
Recent advances in understanding amyotrophic lateral sclerosis and emerging therapies.Fac Rev. 2020 Nov 17;9:12. doi: 10.12703/b/9-12. eCollection 2020. Fac Rev. 2020. PMID: 33659944 Free PMC article. Review.
-
Nuclear import receptors are recruited by FG-nucleoporins to rescue hallmarks of TDP-43 proteinopathy.Mol Neurodegener. 2022 Dec 8;17(1):80. doi: 10.1186/s13024-022-00585-1. Mol Neurodegener. 2022. PMID: 36482422 Free PMC article.
-
Enzymatic and synthetic regulation of polypeptide folding.Chem Sci. 2024 Jan 26;15(7):2282-2299. doi: 10.1039/d3sc05781j. eCollection 2024 Feb 14. Chem Sci. 2024. PMID: 38362427 Free PMC article. Review.
-
Modulating biomolecular condensates: a novel approach to drug discovery.Nat Rev Drug Discov. 2022 Nov;21(11):841-862. doi: 10.1038/s41573-022-00505-4. Epub 2022 Aug 16. Nat Rev Drug Discov. 2022. PMID: 35974095 Free PMC article. Review.
-
Balanced between order and disorder: a new phase in transcription elongation control and beyond.Transcription. 2019 Jun;10(3):157-163. doi: 10.1080/21541264.2019.1570812. Epub 2019 Jan 31. Transcription. 2019. PMID: 30663929 Free PMC article. Review.
References
-
- Brangwynne CP, Tompa P, Pappu RV. Polymer physics of intracellular phase transitions. Nat Phys. 2015;11:899–904.
-
- Brautigam CA. Calculations and Publication-Quality Illustrations for Analytical Ultracentrifugation Data. Methods Enzymol. 2015;562:109–133. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM114274/GM/NIGMS NIH HHS/United States
- S10 OD018027/OD/NIH HHS/United States
- R01 GM083960/GM/NIGMS NIH HHS/United States
- P41 RR001209/RR/NCRR NIH HHS/United States
- R01 GM112108/GM/NIGMS NIH HHS/United States
- T32 GM008297/GM/NIGMS NIH HHS/United States
- R01 GM118530/GM/NIGMS NIH HHS/United States
- T32 GM008203/GM/NIGMS NIH HHS/United States
- S10 RR026461/RR/NCRR NIH HHS/United States
- P41 GM103393/GM/NIGMS NIH HHS/United States
- P41 GM109824/GM/NIGMS NIH HHS/United States
- S10 OD021596/OD/NIH HHS/United States
- R01 GM069909/GM/NIGMS NIH HHS/United States
- U01 GM098256/GM/NIGMS NIH HHS/United States
- R01 GM056322/GM/NIGMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
