

GPR68 Senses Flow and Is Essential for Vascular Physiology
- PMID: 29677517
- PMCID: PMC5951615
- DOI: 10.1016/j.cell.2018.03.076
GPR68 Senses Flow and Is Essential for Vascular Physiology
Abstract
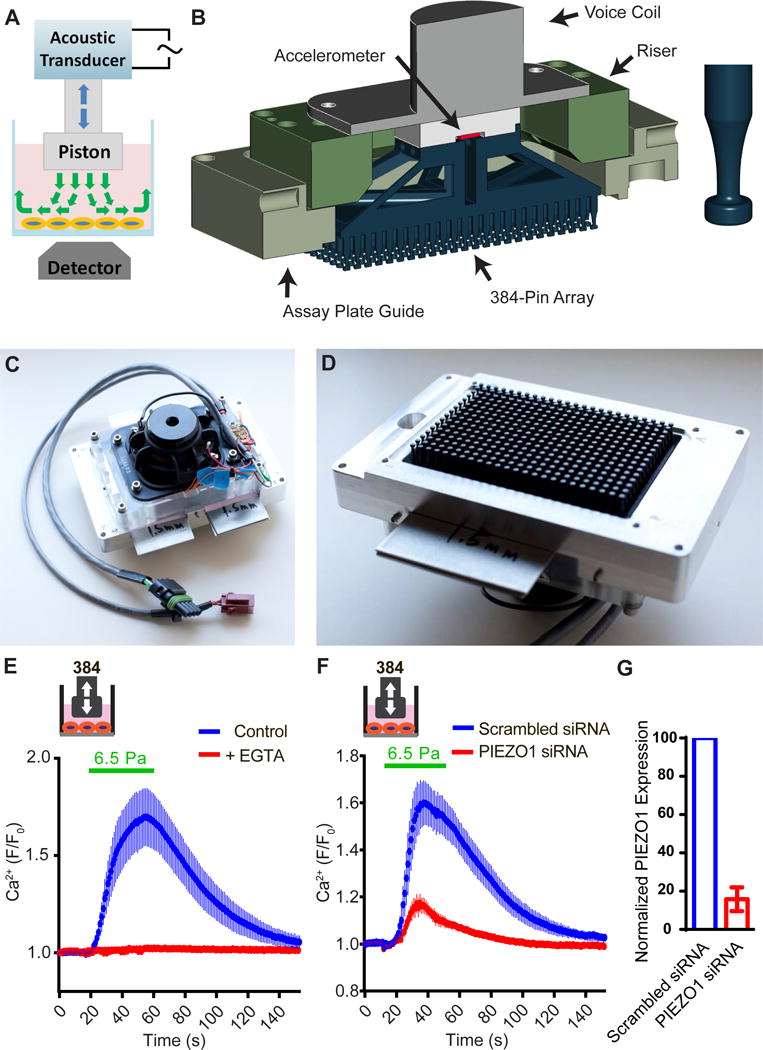
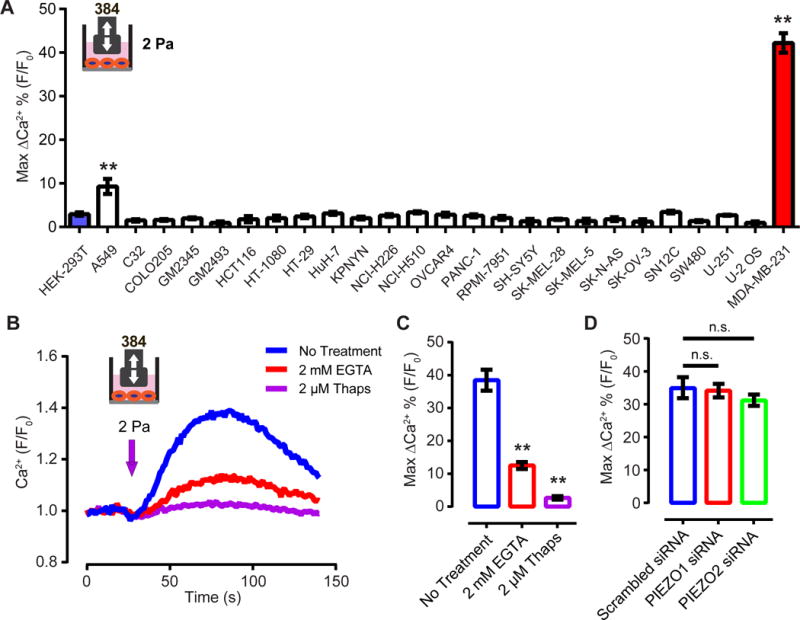
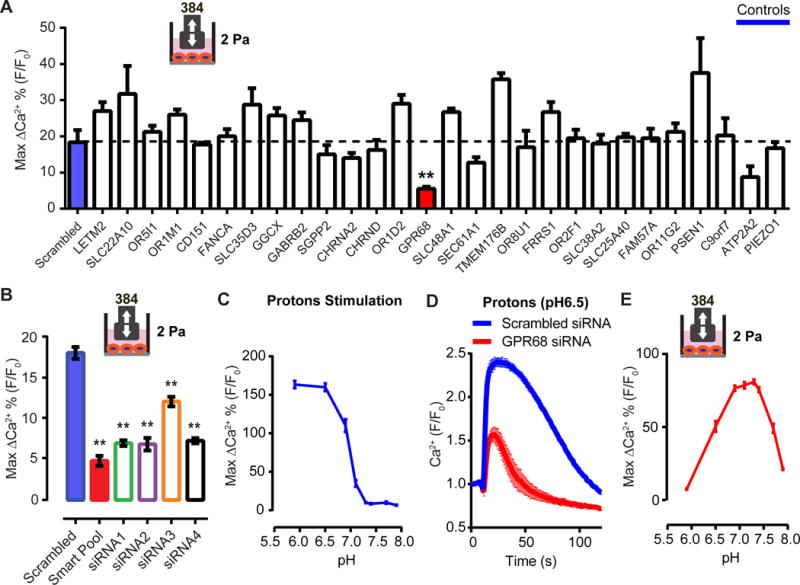
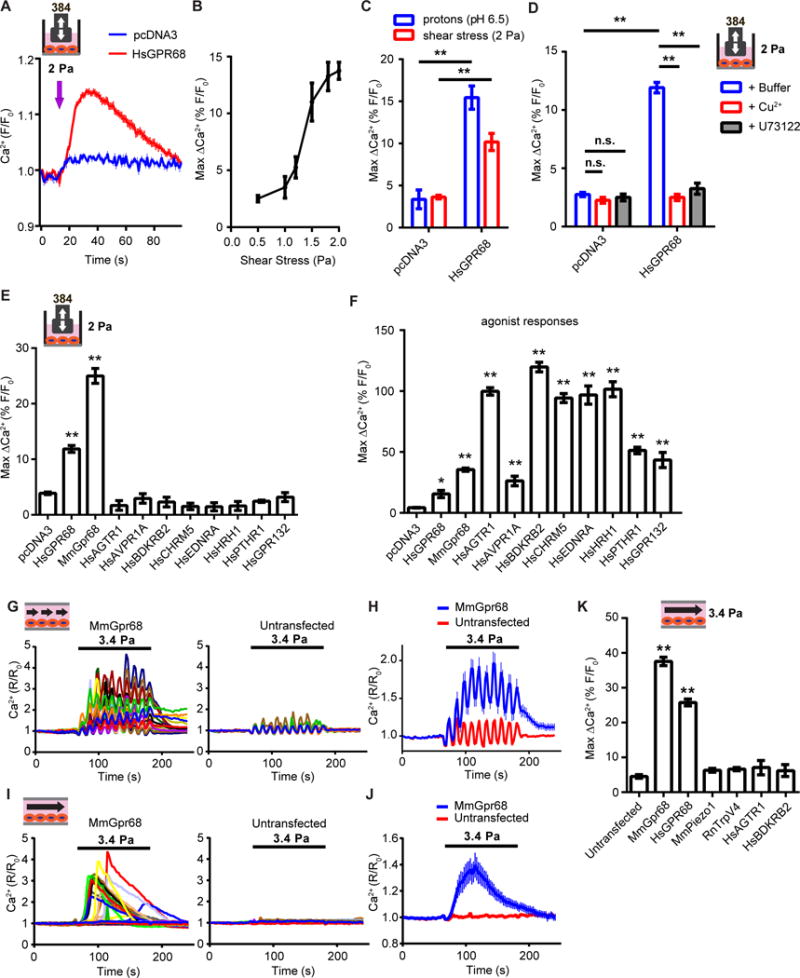
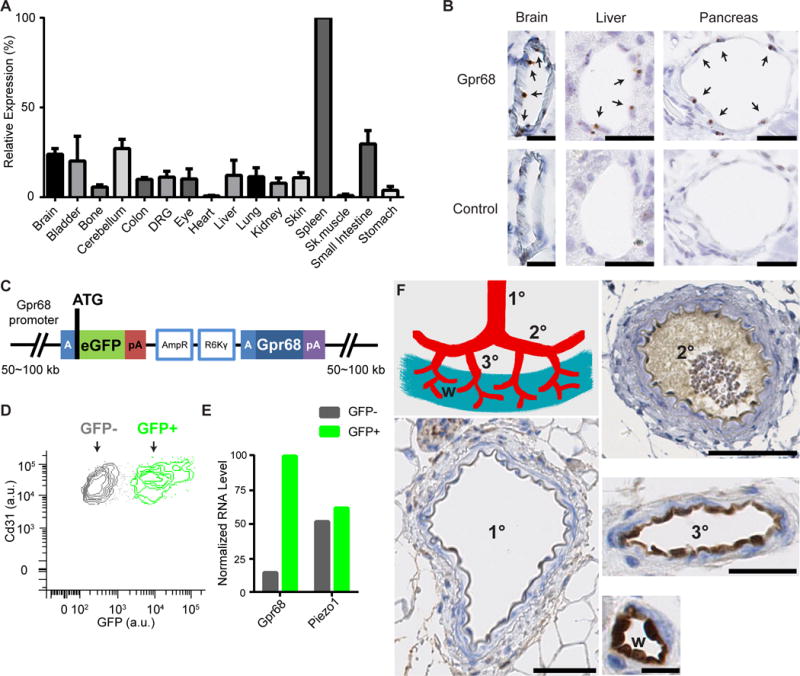
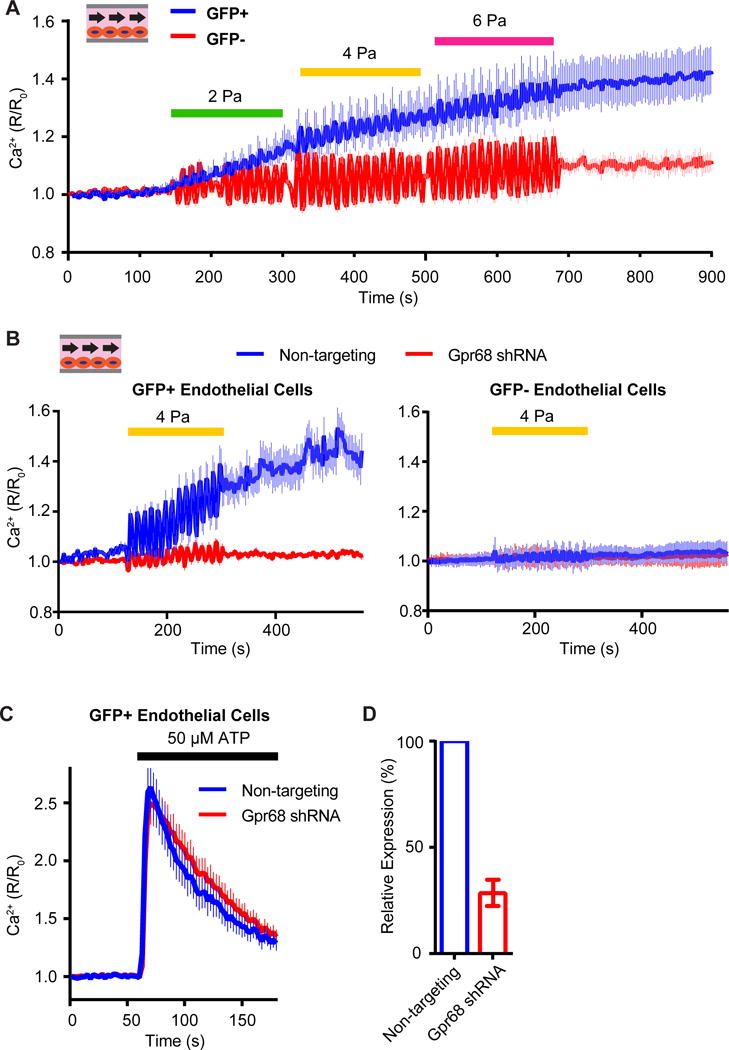
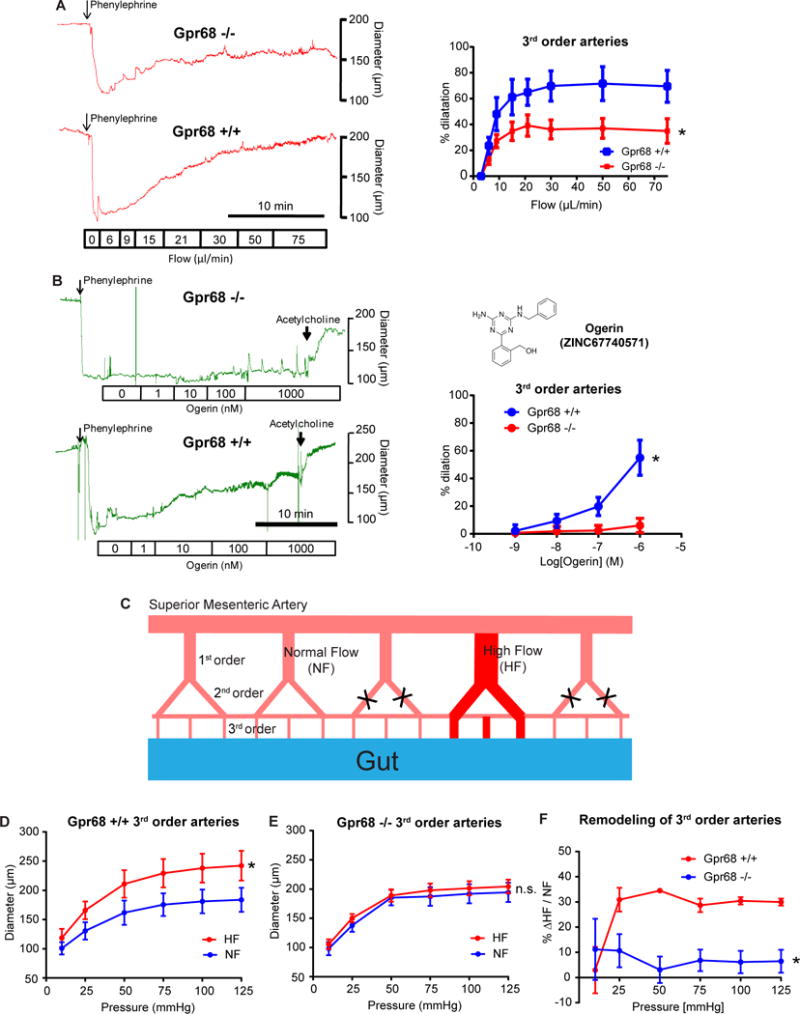
Mechanotransduction plays a crucial role in vascular biology. One example of this is the local regulation of vascular resistance via flow-mediated dilation (FMD). Impairment of this process is a hallmark of endothelial dysfunction and a precursor to a wide array of vascular diseases, such as hypertension and atherosclerosis. Yet the molecules responsible for sensing flow (shear stress) within endothelial cells remain largely unknown. We designed a 384-well screening system that applies shear stress on cultured cells. We identified a mechanosensitive cell line that exhibits shear stress-activated calcium transients, screened a focused RNAi library, and identified GPR68 as necessary and sufficient for shear stress responses. GPR68 is expressed in endothelial cells of small-diameter (resistance) arteries. Importantly, Gpr68-deficient mice display markedly impaired acute FMD and chronic flow-mediated outward remodeling in mesenteric arterioles. Therefore, GPR68 is an essential flow sensor in arteriolar endothelium and is a critical signaling component in cardiovascular pathophysiology.
Keywords: GPCR; blood flow; mechanosensation; mechanotransduction; outward remodeling; shear stress; vascular biology; vasodilation.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
A Mechanosensitive GPCR that Detects the Bloody Force.Cell. 2018 Apr 19;173(3):542-544. doi: 10.1016/j.cell.2018.04.001. Cell. 2018. PMID: 29677505 Free PMC article.
-
Shear elegance: A novel screen uncovers a mechanosensitive GPCR.J Gen Physiol. 2018 Jul 2;150(7):907-910. doi: 10.1085/jgp.201812101. Epub 2018 Jun 14. J Gen Physiol. 2018. PMID: 29903977 Free PMC article.
References
-
- Caillon A, Grenier C, Grimaud L, Vessieres E, Guihot AL, Blanchard S, Lelievre E, Chabbert M, Foucher ED, Jeannin P, et al. The angiotensin II type 2 receptor activates flow-mediated outward remodelling through T cells-dependent interleukin-17 production. Cardiovascular research. 2016;112:515–525. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
