

Recent advances in flower color variation and patterning of Japanese morning glory and petunia
- PMID: 29681755
- PMCID: PMC5903981
- DOI: 10.1270/jsbbs.17107
Recent advances in flower color variation and patterning of Japanese morning glory and petunia
Abstract
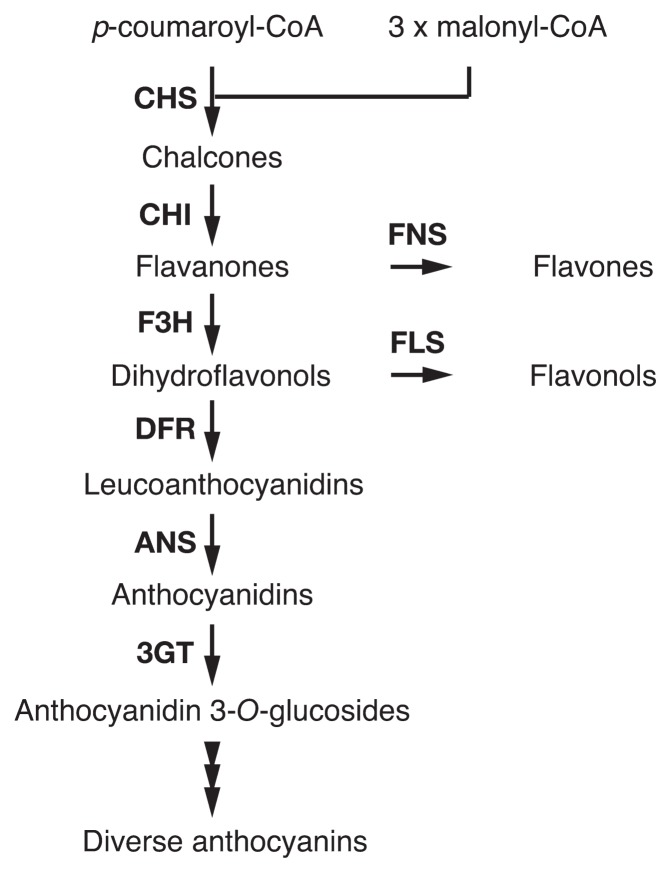
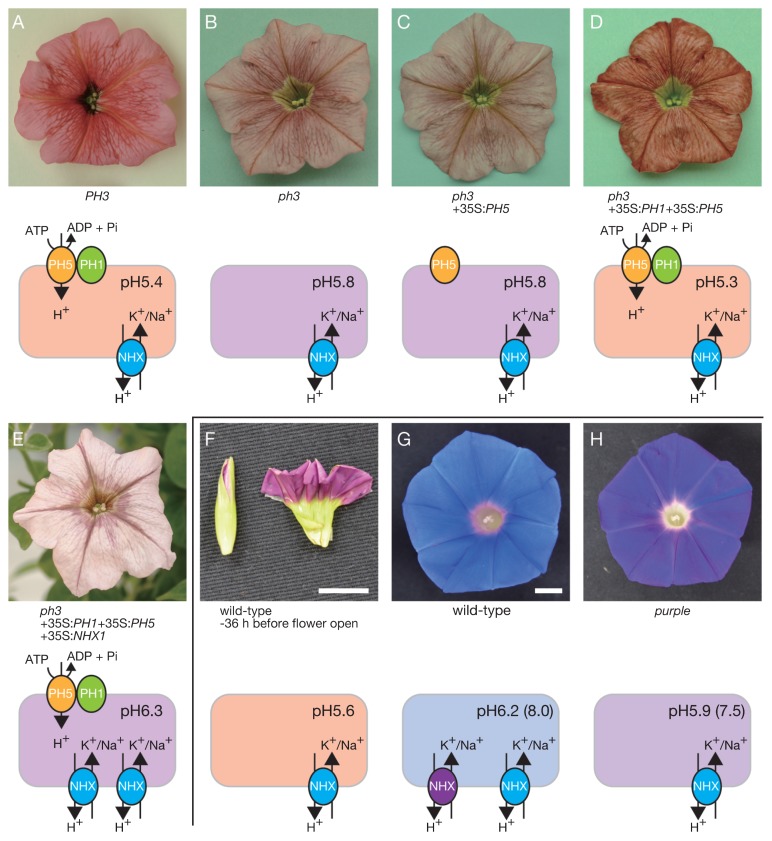
The Japanese morning glory (Ipomoea nil) and petunia (Petunia hybrida), locally called "Asagao" and "Tsukubane-asagao", respectively, are popular garden plants. They have been utilized as model plants for studying the genetic basis of floricultural traits, especially anthocyanin pigmentation in flower petals. In their long history of genetic studies, many mutations affecting flower pigmentation have been characterized, and both structural and regulatory genes for the anthocyanin biosynthesis pathway have been identified. In this review, we will summarize recent advances in the understanding of flower pigmentation in the two species with respect to flower hue and color patterning. Regarding flower hue, we will describe a novel enhancer of flavonoid production that controls the intensity of flower pigmentation, new aspects related to a flavonoid glucosyltransferase that has been known for a long time, and the regulatory mechanisms of vacuolar pH being a key determinant of red and blue coloration. On color patterning, we describe particular flower patterns regulated by epigenetic and RNA-silencing mechanisms. As high-quality whole genome sequences of the Japanese morning glory and petunia wild parents (P. axillaris and P. inflata, respectively) were published in 2016, further study on flower pigmentation will be accelerated.
Keywords: Ipomoea; RNA silencing; anthocyanin; epigenetics; floral pigmentation pattern; petunia; vacuolar pH.
Figures

Similar articles
-
Genetics and epigenetics in flower pigmentation associated with transposable elements in morning glories.Adv Biophys. 2004;38:141-59. Adv Biophys. 2004. PMID: 15493332 Review.
-
A chalcone isomerase-like protein enhances flavonoid production and flower pigmentation.Plant J. 2014 Apr;78(2):294-304. doi: 10.1111/tpj.12469. Epub 2014 Mar 14. Plant J. 2014. PMID: 24517863
-
Genes encoding the vacuolar Na+/H+ exchanger and flower coloration.Plant Cell Physiol. 2001 May;42(5):451-61. doi: 10.1093/pcp/pce080. Plant Cell Physiol. 2001. PMID: 11382810
-
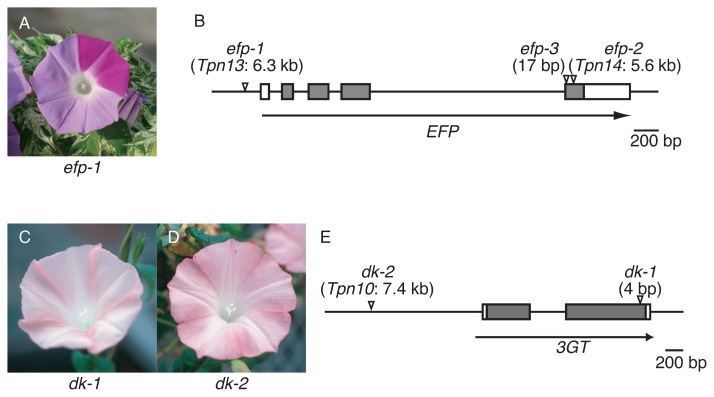
Japanese morning glory dusky mutants displaying reddish-brown or purplish-gray flowers are deficient in a novel glycosylation enzyme for anthocyanin biosynthesis, UDP-glucose:anthocyanidin 3-O-glucoside-2''-O-glucosyltransferase, due to 4-bp insertions in the gene.Plant J. 2005 May;42(3):353-63. doi: 10.1111/j.1365-313X.2005.02383.x. Plant J. 2005. PMID: 15842621
-
Achievements and perspectives in biochemistry concerning anthocyanin modification for blue flower coloration.Plant Cell Physiol. 2015 Jan;56(1):28-40. doi: 10.1093/pcp/pcu097. Epub 2014 Jul 10. Plant Cell Physiol. 2015. PMID: 25015943 Review.
Cited by
-
Integrative Analysis of Metabolome and Transcriptome Reveals the Mechanism of Color Formation in Yellow-Fleshed Kiwifruit.Int J Mol Sci. 2023 Jan 13;24(2):1573. doi: 10.3390/ijms24021573. Int J Mol Sci. 2023. PMID: 36675098 Free PMC article.
-
Wild and weedy Hesperis matronalis hosts turnip mosaic virus across heterogeneous landscapes in upstate New York.Virus Res. 2023 Jan 2;323:199011. doi: 10.1016/j.virusres.2022.199011. Epub 2022 Nov 28. Virus Res. 2023. PMID: 36511291 Free PMC article.
-
Unveiling the Petunia hybrida Virome: Metatranscriptomic Profiling from the Bulgarian Market and In Vitro Cultures.Plants (Basel). 2025 Aug 21;14(16):2597. doi: 10.3390/plants14162597. Plants (Basel). 2025. PMID: 40872220 Free PMC article.
-
Genome-wide comparative analysis of the nucleotide-binding site-encoding genes in four Ipomoea species.Front Plant Sci. 2022 Aug 18;13:960723. doi: 10.3389/fpls.2022.960723. eCollection 2022. Front Plant Sci. 2022. PMID: 36061812 Free PMC article.
-
Phylogenetic Analysis of the SQUAMOSA Promoter-Binding Protein-Like Genes in Four Ipomoea Species and Expression Profiling of the IbSPLs During Storage Root Development in Sweet Potato (Ipomoea batatas).Front Plant Sci. 2022 Jan 21;12:801061. doi: 10.3389/fpls.2021.801061. eCollection 2021. Front Plant Sci. 2022. PMID: 35126426 Free PMC article.
References
-
- Appelhagen, I., Nordholt, N., Seidel, T., Spelt, K., Koes, R., Quattrochio, F., Sagasser, M. and Weisshaar, B. (2015) TRANSPARENT TESTA 13 is a tonoplast P3A-ATPase required for vacuolar deposition of proanthocyanidins in Arabidopsis thaliana seeds. Plant J. 82: 840–849. - PubMed
-
- Bassil, E., Tajima, H., Liang, Y.C., Ohto, M.A., Ushijima, K., Nakano, R., Esumi, T., Coku, A., Belmonte, M. and Blumwald, E. (2011) The Arabidopsis Na+/H+ antiporters NHX1 and NHX2 control vacuolar pH and K+ homeostasis to regulate growth, flower development, and reproduction. Plant Cell 23: 3482–3497. - PMC - PubMed
-
- Bassil, E. and Blumwald, E. (2014) The ins and outs of intracellular ion homeostasis: NHX-type cation/H+ transporters. Curr. Opin. Plant Biol. 22: 1–6. - PubMed
-
- Bombarely, A., Moser, M., Amrad, A., Bao, M., Bapaume, L., Barry, C.S., Bliek, M., Boersma, M.R., Borghi, L., Bruggmann, R. et al. (2016) Insight into the evolution of the Solanaceae from the parental genomes of Petunia hybrida. Nat. Plants 2: 16074. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials