

Switchable slow cellular conductances determine robustness and tunability of network states
- PMID: 29684009
- PMCID: PMC5940245
- DOI: 10.1371/journal.pcbi.1006125
Switchable slow cellular conductances determine robustness and tunability of network states
Abstract
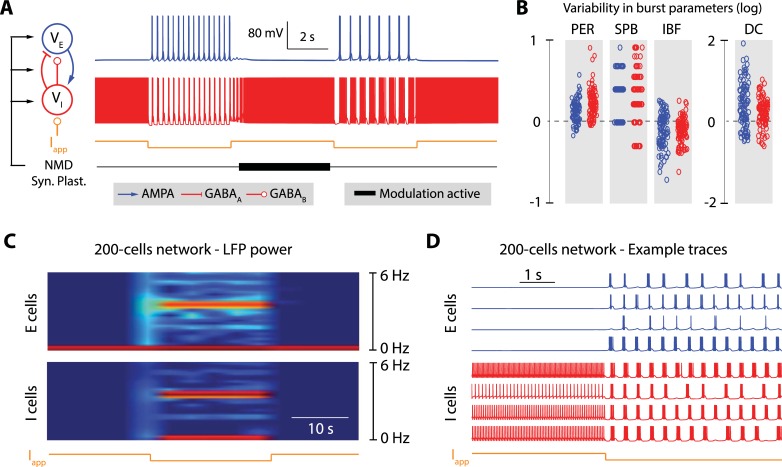
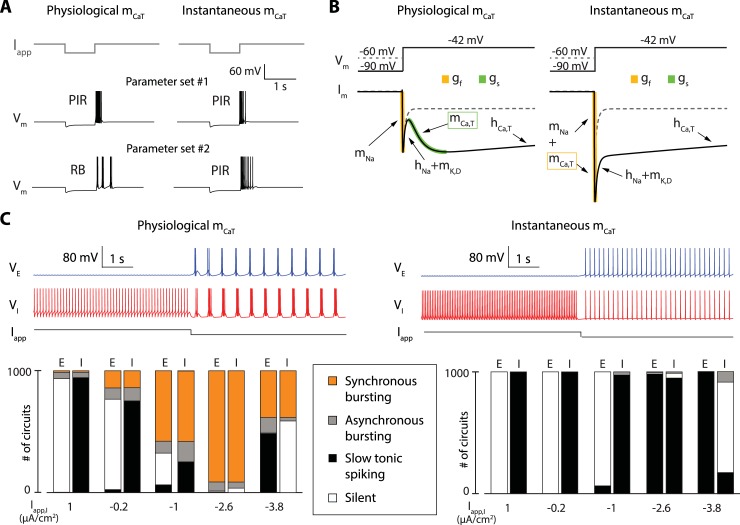
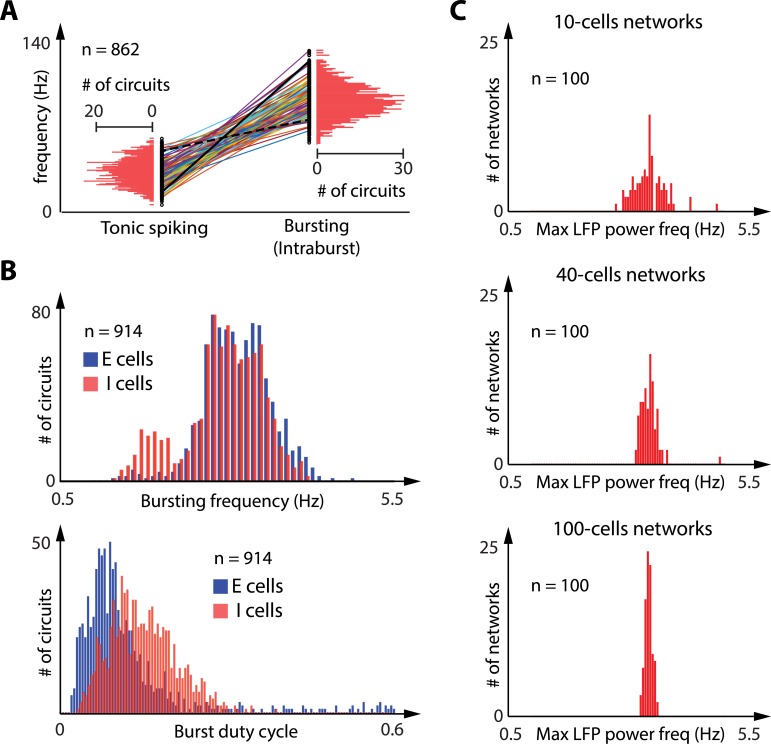
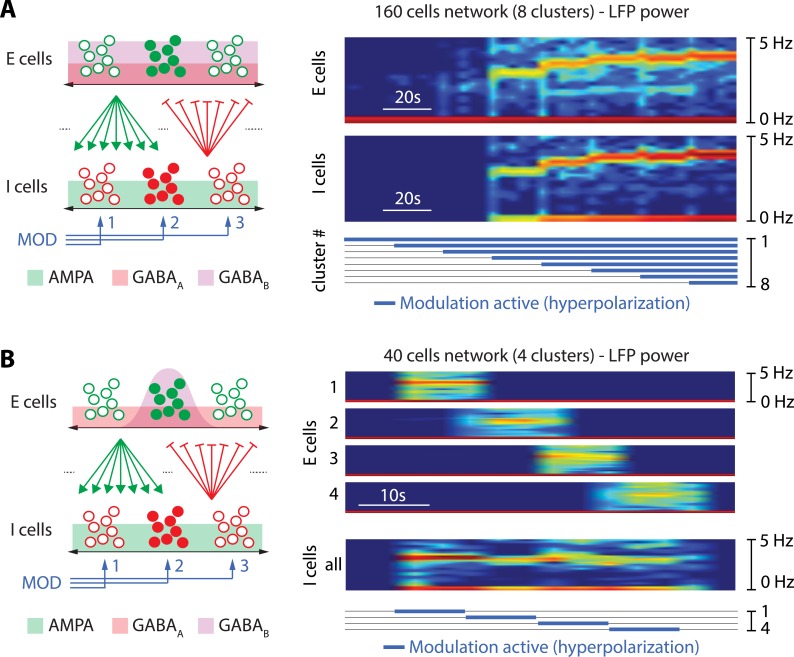
Neuronal information processing is regulated by fast and localized fluctuations of brain states. Brain states reliably switch between distinct spatiotemporal signatures at a network scale even though they are composed of heterogeneous and variable rhythms at a cellular scale. We investigated the mechanisms of this network control in a conductance-based population model that reliably switches between active and oscillatory mean-fields. Robust control of the mean-field properties relies critically on a switchable negative intrinsic conductance at the cellular level. This conductance endows circuits with a shared cellular positive feedback that can switch population rhythms on and off at a cellular resolution. The switch is largely independent from other intrinsic neuronal properties, network size and synaptic connectivity. It is therefore compatible with the temporal variability and spatial heterogeneity induced by slower regulatory functions such as neuromodulation, synaptic plasticity and homeostasis. Strikingly, the required cellular mechanism is available in all cell types that possess T-type calcium channels but unavailable in computational models that neglect the slow kinetics of their activation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Robust switches in thalamic network activity require a timescale separation between sodium and T-type calcium channel activations.PLoS Comput Biol. 2021 May 18;17(5):e1008997. doi: 10.1371/journal.pcbi.1008997. eCollection 2021 May. PLoS Comput Biol. 2021. PMID: 34003841 Free PMC article.
-
Cellular switches orchestrate rhythmic circuits.Biol Cybern. 2019 Apr;113(1-2):71-82. doi: 10.1007/s00422-018-0778-6. Epub 2018 Sep 3. Biol Cybern. 2019. PMID: 30178150
-
Effects of cellular homeostatic intrinsic plasticity on dynamical and computational properties of biological recurrent neural networks.J Neurosci. 2013 Sep 18;33(38):15032-43. doi: 10.1523/JNEUROSCI.0870-13.2013. J Neurosci. 2013. PMID: 24048833 Free PMC article.
-
Plasticity in single neuron and circuit computations.Nature. 2004 Oct 14;431(7010):789-95. doi: 10.1038/nature03011. Nature. 2004. PMID: 15483600 Review.
-
Synaptic plasticity: taming the beast.Nat Neurosci. 2000 Nov;3 Suppl:1178-83. doi: 10.1038/81453. Nat Neurosci. 2000. PMID: 11127835 Review.
Cited by
-
Co-opting evo-devo concepts for new insights into mechanisms of behavioural diversity.J Exp Biol. 2019 Apr 15;222(Pt 8):jeb190058. doi: 10.1242/jeb.190058. J Exp Biol. 2019. PMID: 30988051 Free PMC article. Review.
-
Robust switches in thalamic network activity require a timescale separation between sodium and T-type calcium channel activations.PLoS Comput Biol. 2021 May 18;17(5):e1008997. doi: 10.1371/journal.pcbi.1008997. eCollection 2021 May. PLoS Comput Biol. 2021. PMID: 34003841 Free PMC article.
References
-
- Zagha E, McCormick DA (2014) Neural control of brain state. Curr Opin Neurobiol. 29:178–86. doi: 10.1016/j.conb.2014.09.010 - DOI - PMC - PubMed
-
- Buzsaki G (2006) Rhythms of the brain New York: Oxford University Press.
-
- Wang XJ (2010) Neurophysiological and computational principles of cortical rhythms in cognition. Physiol Rev. 90:1195–268. doi: 10.1152/physrev.00035.2008 - DOI - PMC - PubMed
-
- Uhlhaas PJ, Singer W (2006) Neural synchrony in brain disorders: relevance for cognitive dysfunctions and pathophysiology. Neuron. 52:155–68. doi: 10.1016/j.neuron.2006.09.020 - DOI - PubMed
-
- Lee SH, Dan Y (2012) Neuromodulation of brain states. Neuron. 76:209–22. doi: 10.1016/j.neuron.2012.09.012 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
