

PgaB orthologues contain a glycoside hydrolase domain that cleaves deacetylated poly-β(1,6)-N-acetylglucosamine and can disrupt bacterial biofilms
- PMID: 29684093
- PMCID: PMC5933820
- DOI: 10.1371/journal.ppat.1006998
PgaB orthologues contain a glycoside hydrolase domain that cleaves deacetylated poly-β(1,6)-N-acetylglucosamine and can disrupt bacterial biofilms
Abstract
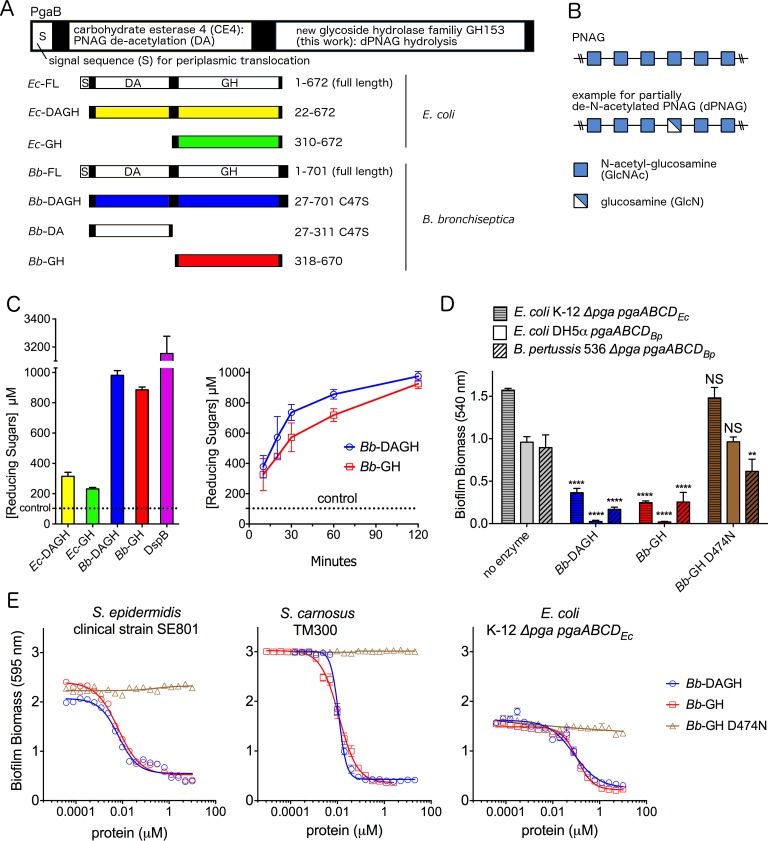
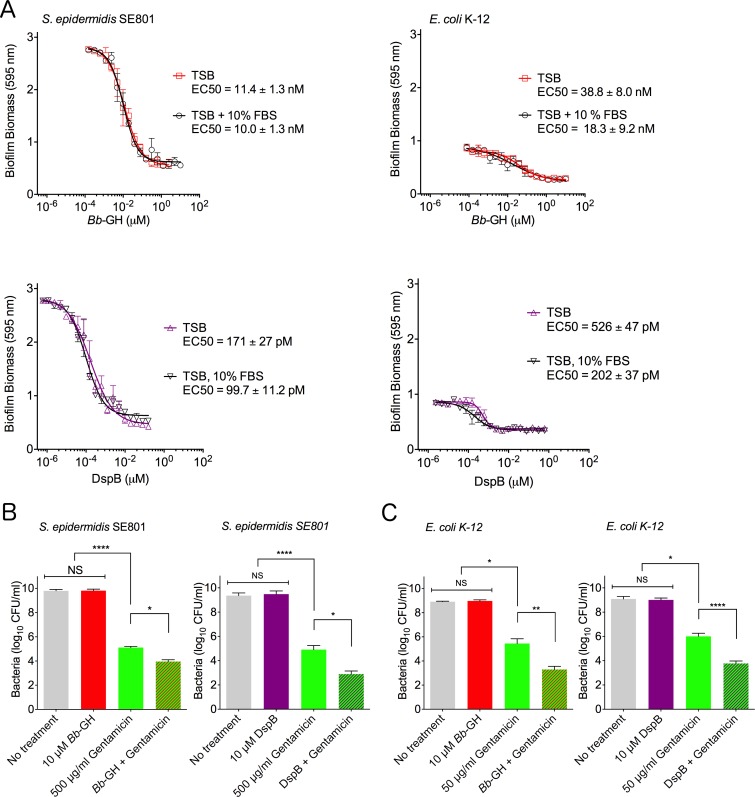
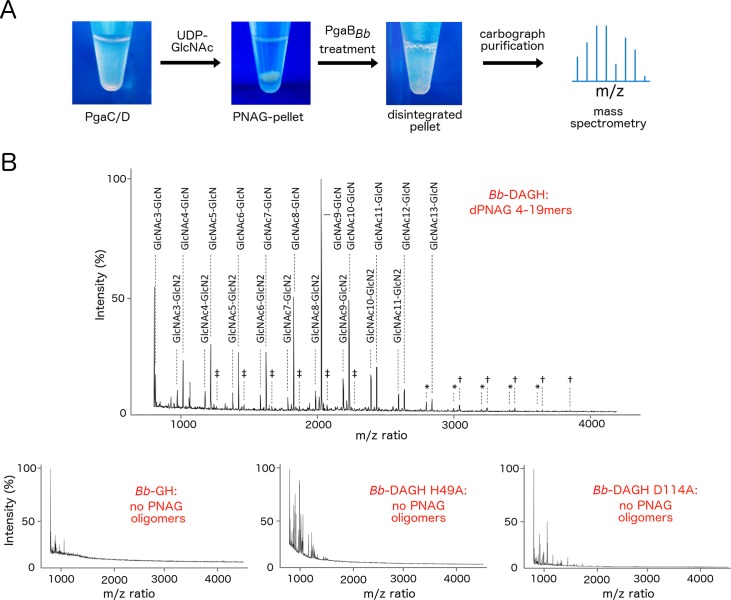
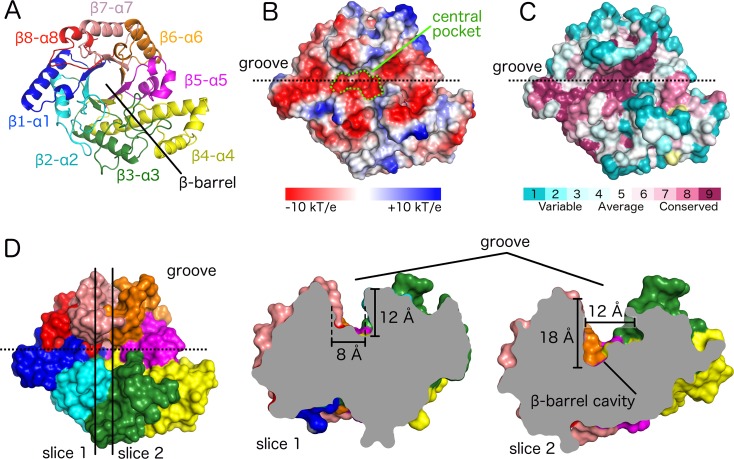
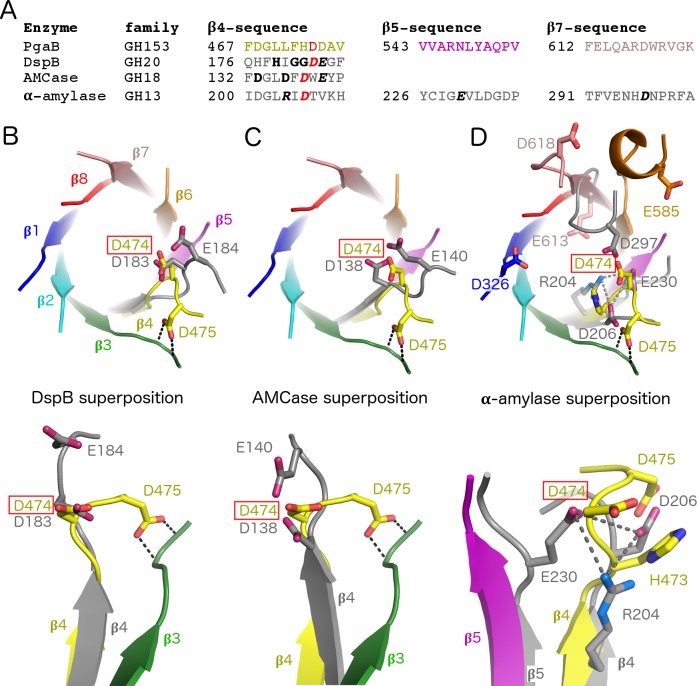
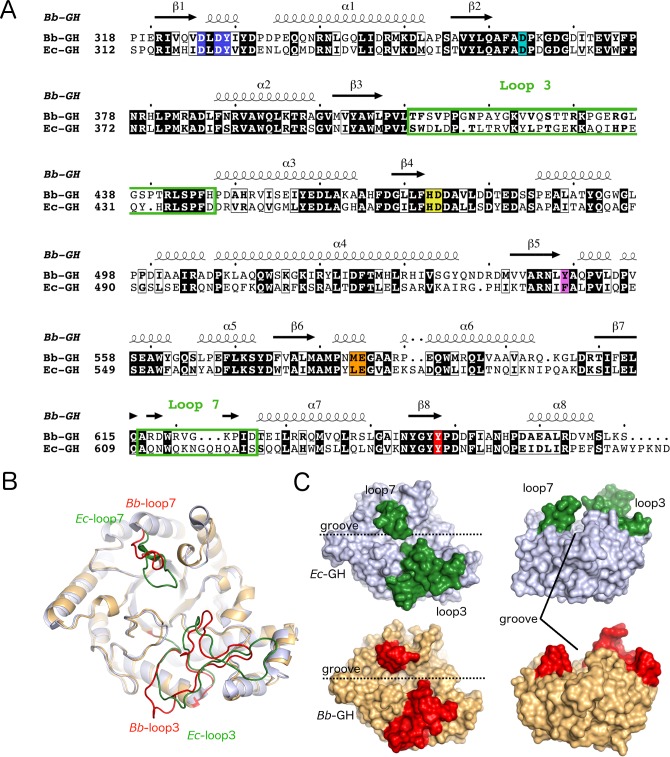
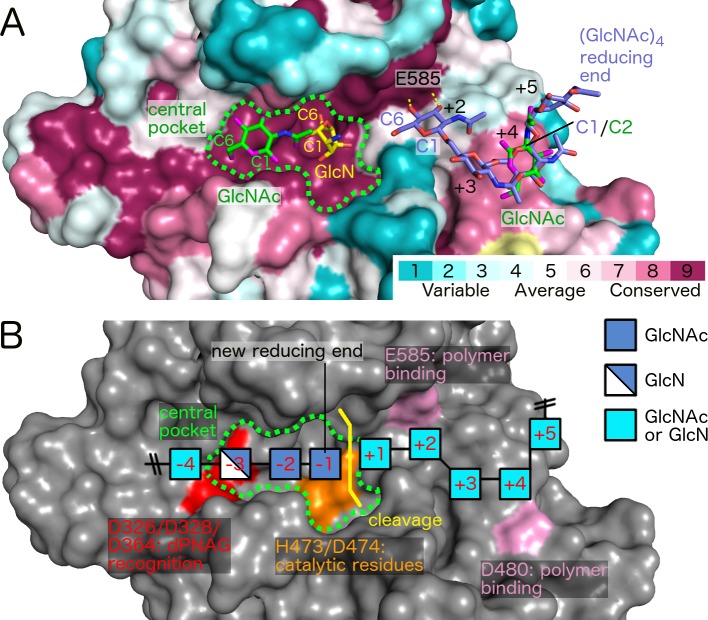
Poly-β(1,6)-N-acetyl-D-glucosamine (PNAG) is a major biofilm component of many pathogenic bacteria. The production, modification, and export of PNAG in Escherichia coli and Bordetella species require the protein products encoded by the pgaABCD operon. PgaB is a two-domain periplasmic protein that contains an N-terminal deacetylase domain and a C-terminal PNAG binding domain that is critical for export. However, the exact function of the PgaB C-terminal domain remains unclear. Herein, we show that the C-terminal domains of Bordetella bronchiseptica PgaB (PgaBBb) and E. coli PgaB (PgaBEc) function as glycoside hydrolases. These enzymes hydrolyze purified deacetylated PNAG (dPNAG) from Staphylococcus aureus, disrupt PNAG-dependent biofilms formed by Bordetella pertussis, Staphylococcus carnosus, Staphylococcus epidermidis, and E. coli, and potentiate bacterial killing by gentamicin. Furthermore, we found that PgaBBb was only able to hydrolyze PNAG produced in situ by the E. coli PgaCD synthase complex when an active deacetylase domain was present. Mass spectrometry analysis of the PgaB-hydrolyzed dPNAG substrate showed a GlcN-GlcNAc-GlcNAc motif at the new reducing end of detected fragments. Our 1.76 Å structure of the C-terminal domain of PgaBBb reveals a central cavity within an elongated surface groove that appears ideally suited to recognize the GlcN-GlcNAc-GlcNAc motif. The structure, in conjunction with molecular modeling and site directed mutagenesis led to the identification of the dPNAG binding subsites and D474 as the probable catalytic acid. This work expands the role of PgaB within the PNAG biosynthesis machinery, defines a new glycoside hydrolase family GH153, and identifies PgaB as a possible therapeutic agent for treating PNAG-dependent biofilm infections.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: A patent covering the use of the PgaB and other glycoside hydrolases for the prevention and/or disruption of biofilms has been filed (Application number WO2015/184526 A1).
Figures


References
-
- Mishra M, Parise G, Jackson KD, Wozniak DJ, Deora R. The BvgAS signal transduction system regulates biofilm development in Bordetella. J Bacteriol. 2005;187(4):1474–84. doi: 10.1128/JB.187.4.1474-1484.2005 ; PubMed Central PMCID: PMC545624. - DOI - PMC - PubMed
-
- Irie Y, Mattoo S, Yuk MH. The Bvg virulence control system regulates biofilm formation in Bordetella bronchiseptica. J Bacteriol. 2004;186(17):5692–8. doi: 10.1128/JB.186.17.5692-5698.2004 ; PubMed Central PMCID: PMC516841. - DOI - PMC - PubMed
-
- Sukumar N, Nicholson TL, Conover MS, Ganguly T, Deora R. Comparative analyses of a cystic fibrosis isolate of Bordetella bronchiseptica reveal differences in important pathogenic phenotypes. Infect Immun. 2014;82(4):1627–37. doi: 10.1128/IAI.01453-13 ; PubMed Central PMCID: PMC3993408. - DOI - PMC - PubMed
-
- Serra DO, Conover MS, Arnal L, Sloan GP, Rodriguez ME, Yantorno OM, et al. FHA-mediated cell-substrate and cell-cell adhesions are critical for Bordetella pertussis biofilm formation on abiotic surfaces and in the mouse nose and the trachea. PLoS One. 2011;6(12):e28811 Epub 2012/01/05. doi: 10.1371/journal.pone.0028811 PONE-D-11-15276 [pii]. ; PubMed Central PMCID: PMC3245231. - DOI - PMC - PubMed
-
- Conover MS, Mishra M, Deora R. Extracellular DNA is essential for maintaining Bordetella biofilm integrity on abiotic surfaces and in the upper respiratory tract of mice. PLoS One. 2011;6(2):e16861 Epub 2011/02/25. doi: 10.1371/journal.pone.0016861 . - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
