

The past and presence of gene targeting: from chemicals and DNA via proteins to RNA
- PMID: 29685979
- PMCID: PMC5915719
- DOI: 10.1098/rstb.2017.0077
The past and presence of gene targeting: from chemicals and DNA via proteins to RNA
Abstract
The ability to target DNA specifically at any given position within the genome allows many intriguing possibilities and has inspired scientists for decades. Early gene-targeting efforts exploited chemicals or DNA oligonucleotides to interfere with the DNA at a given location in order to inactivate a gene or to correct mutations. We here describe an example towards correcting a genetic mutation underlying Pompe's disease using a nucleotide-fused nuclease (TFO-MunI). In addition to the promise of gene correction, scientists soon realized that genes could be inactivated or even re-activated without inducing potentially harmful DNA damage by targeting transcriptional modulators to a particular gene. However, it proved difficult to fuse protein effector domains to the first generation of programmable DNA-binding agents. The engineering of gene-targeting proteins (zinc finger proteins (ZFPs), transcription activator-like effectors (TALEs)) circumvented this problem. The disadvantage of protein-based gene targeting is that a fusion protein needs to be engineered for every locus. The recent introduction of CRISPR/Cas offers a flexible approach to target a (fusion) protein to the locus of interest using cheap designer RNA molecules. Many research groups now exploit this platform and the first human clinical trials have been initiated: CRISPR/Cas has kicked off a new era of gene targeting and is revolutionizing biomedical sciences.This article is part of a discussion meeting issue 'Frontiers in epigenetic chemical biology'.
Keywords: CRISPR/dCas; TALEs (transcription activator-like effectors); ZFPs (zinc finger proteins); genome editing; polyamides.
© 2018 The Author(s).
Conflict of interest statement
We declare we have no competing interests.
Figures
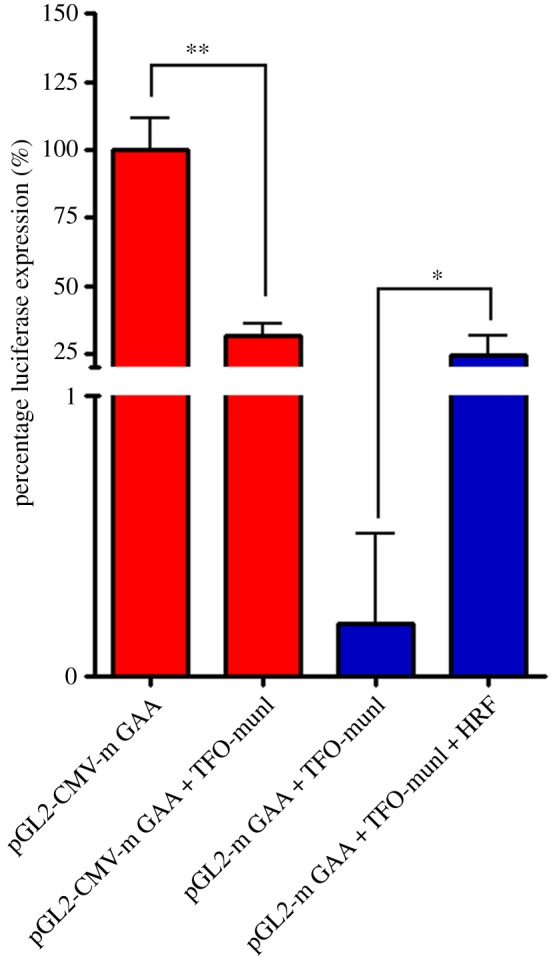
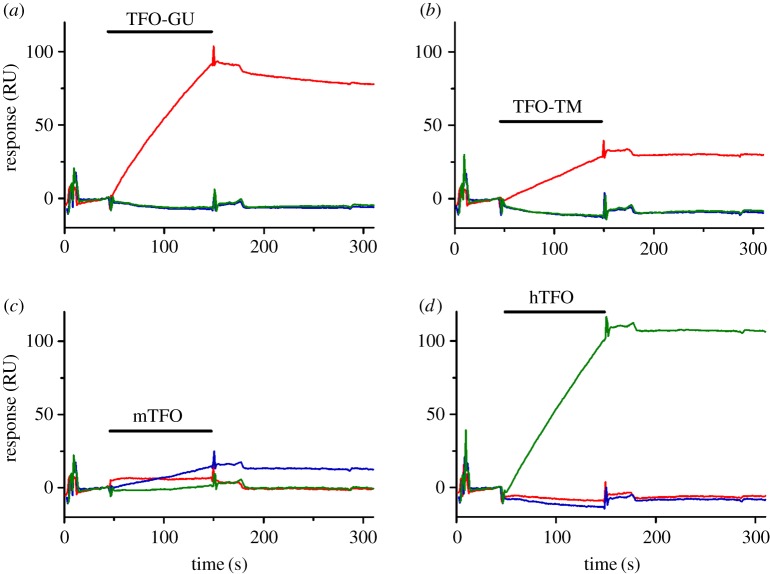
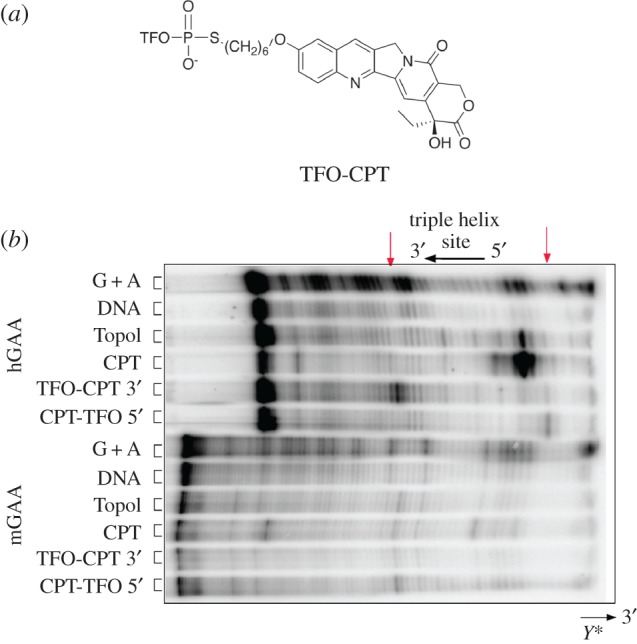
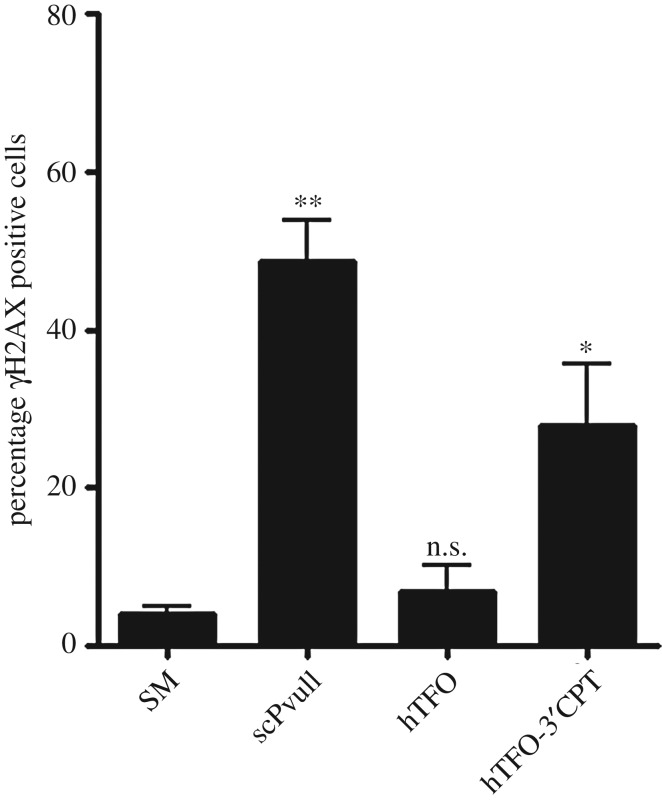
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
