

Characterization of a Two-Component System Transcriptional Regulator, LtdR, That Impacts Group B Streptococcal Colonization and Disease
- PMID: 29685987
- PMCID: PMC6013667
- DOI: 10.1128/IAI.00822-17
Characterization of a Two-Component System Transcriptional Regulator, LtdR, That Impacts Group B Streptococcal Colonization and Disease
Abstract
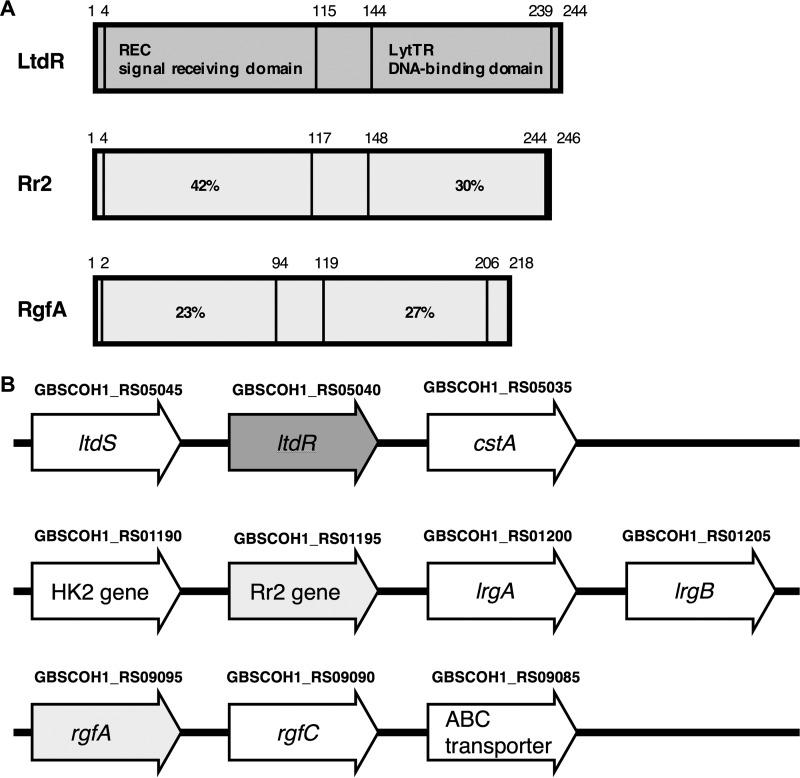
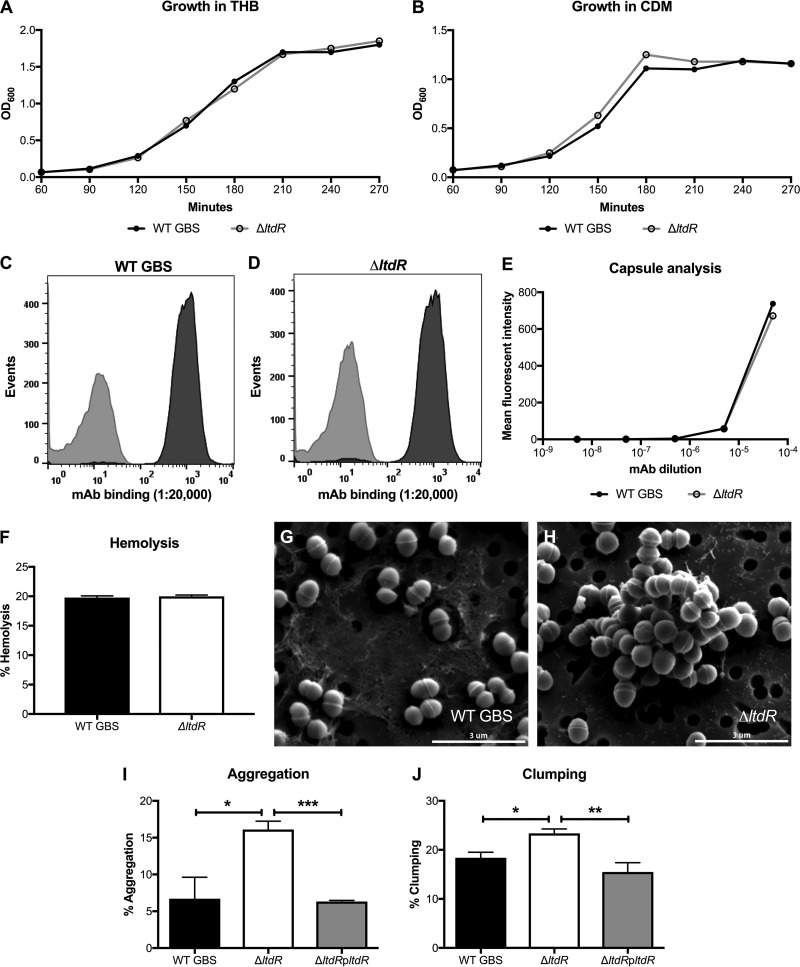
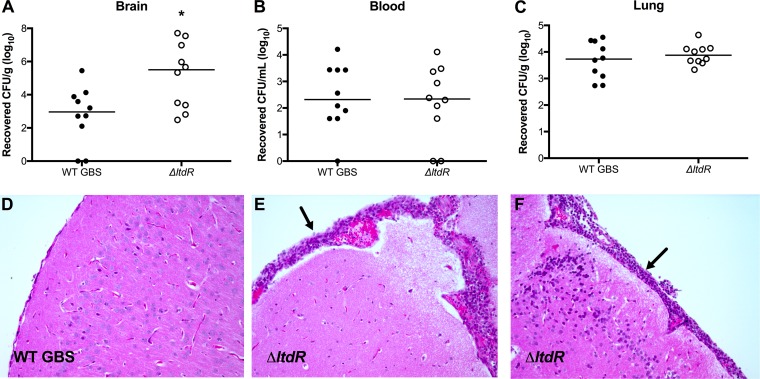
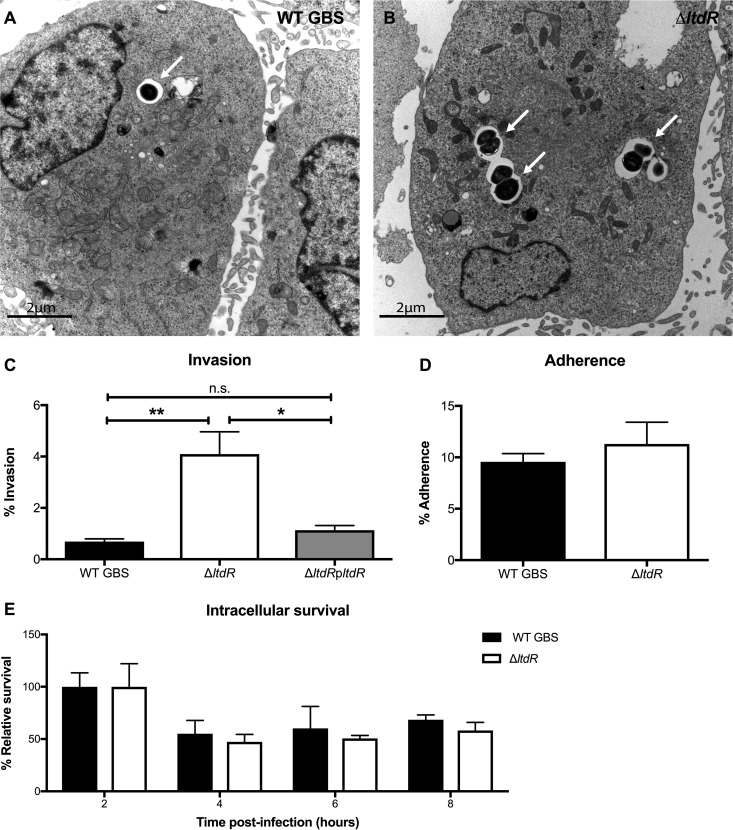
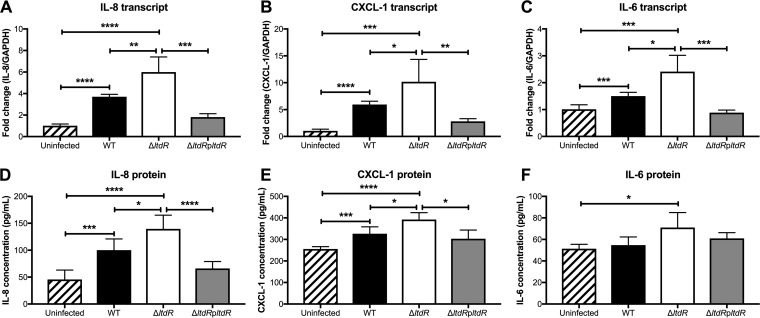
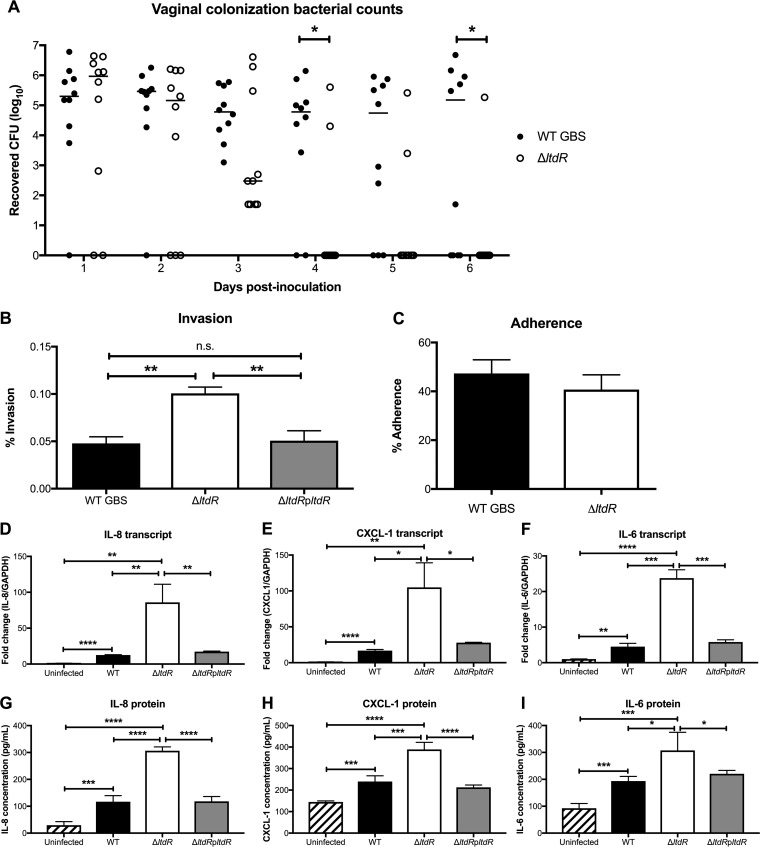
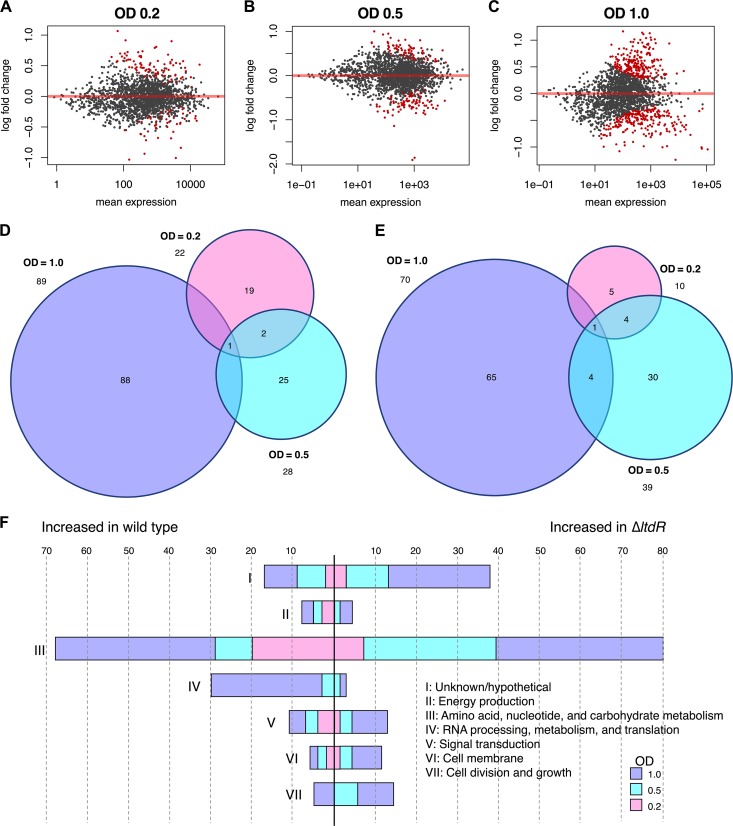
Streptococcus agalactiae (group B Streptococcus [GBS]) is often a commensal bacterium that colonizes healthy adults asymptomatically and is a frequent inhabitant of the vaginal tract in women. However, in immunocompromised individuals, particularly the newborn, GBS may transition to an invasive pathogen and cause serious disease. Despite the use of the currently recommended intrapartum antibiotic prophylaxis for GBS-positive mothers, GBS remains a leading cause of neonatal septicemia and meningitis. To adapt to the various host environments encountered during its disease cycle, GBS possesses multiple two-component regulatory systems (TCSs). Here we investigated the contribution of a transcriptional regulator containing a LytTR domain, LtdR, to GBS pathogenesis. Disruption of the ltdR gene in the GBS chromosome resulted in a significant increase in bacterial invasion into human cerebral microvascular endothelial cells (hCMEC) in vitro as well as the greater penetration of the blood-brain barrier (BBB) and the development of meningitis in vivo Correspondingly, infection of hCMEC with the ΔltdR mutant resulted in increased secretion of the proinflammatory cytokines interleukin-8 (IL-8), CXCL-1, and IL-6. Further, using a mouse model of GBS vaginal colonization, we observed that the ΔltdR mutant was cleared more readily from the vaginal tract and also that infection with the ΔltdR mutant resulted in increased cytokine production from human vaginal epithelial cells. RNA sequencing revealed global transcriptional differences between the ΔltdR mutant and the parental wild-type GBS strain. These results suggest that LtdR regulates many bacterial processes that can influence GBS-host interactions to promote both bacterial persistence and disease progression.
Keywords: RNA sequencing; blood-brain barrier; cytokines; group B Streptococcus; meningitis; two-component regulatory systems; vaginal colonization.
Copyright © 2018 American Society for Microbiology.
Figures
Similar articles
-
Two-Component Signal Transduction Systems in the Human Pathogen Streptococcus agalactiae.Infect Immun. 2020 Jun 22;88(7):e00931-19. doi: 10.1128/IAI.00931-19. Print 2020 Jun 22. Infect Immun. 2020. PMID: 31988177 Free PMC article. Review.
-
The Streptococcus agalactiae LytSR two-component regulatory system promotes vaginal colonization and virulence in vivo.Microbiol Spectr. 2024 Nov 5;12(11):e0197024. doi: 10.1128/spectrum.01970-24. Epub 2024 Oct 14. Microbiol Spectr. 2024. PMID: 39400158 Free PMC article.
-
The Group B Streptococcal Adhesin BspC Interacts with Host Cytokeratin 19 To Promote Colonization of the Female Reproductive Tract.mBio. 2022 Oct 26;13(5):e0178122. doi: 10.1128/mbio.01781-22. Epub 2022 Sep 7. mBio. 2022. PMID: 36069447 Free PMC article.
-
Genomic Analyses Identify Manganese Homeostasis as a Driver of Group B Streptococcal Vaginal Colonization.mBio. 2022 Jun 28;13(3):e0098522. doi: 10.1128/mbio.00985-22. Epub 2022 Jun 6. mBio. 2022. PMID: 35658538 Free PMC article.
-
Group B Streptococcal Maternal Colonization and Neonatal Disease: Molecular Mechanisms and Preventative Approaches.Front Pediatr. 2018 Feb 22;6:27. doi: 10.3389/fped.2018.00027. eCollection 2018. Front Pediatr. 2018. PMID: 29520354 Free PMC article. Review.
Cited by
-
Occludin and collagen IV degradation mediated by the T9SS effector SspA contributes to blood-brain barrier damage in ducks during Riemerella anatipestifer infection.Vet Res. 2024 Apr 9;55(1):49. doi: 10.1186/s13567-024-01304-y. Vet Res. 2024. PMID: 38594770 Free PMC article.
-
Heterogeneity of the group B streptococcal type VII secretion system and influence on colonization of the female genital tract.bioRxiv [Preprint]. 2023 Jan 25:2023.01.25.525443. doi: 10.1101/2023.01.25.525443. bioRxiv. 2023. Update in: Mol Microbiol. 2023 Aug;120(2):258-275. doi: 10.1111/mmi.15115. PMID: 36747681 Free PMC article. Updated. Preprint.
-
In Vivo Role of Two-Component Regulatory Systems in Models of Urinary Tract Infections.Pathogens. 2023 Jan 10;12(1):119. doi: 10.3390/pathogens12010119. Pathogens. 2023. PMID: 36678467 Free PMC article. Review.
-
Two-Component Signal Transduction Systems in the Human Pathogen Streptococcus agalactiae.Infect Immun. 2020 Jun 22;88(7):e00931-19. doi: 10.1128/IAI.00931-19. Print 2020 Jun 22. Infect Immun. 2020. PMID: 31988177 Free PMC article. Review.
-
Group B streptococcal infections in pregnancy and early life.Clin Microbiol Rev. 2025 Mar 13;38(1):e0015422. doi: 10.1128/cmr.00154-22. Epub 2024 Nov 25. Clin Microbiol Rev. 2025. PMID: 39584819 Review.
References
-
- Thigpen MC, Whitney CG, Messonnier NE, Zell ER, Lynfield R, Hadler JL, Harrison LH, Farley MM, Reingold A, Bennett NM, Craig AS, Schaffner W, Thomas A, Lewis MM, Scallan E, Schuchat A, Emerging Infections Programs Network. 2011. Bacterial meningitis in the United States, 1998-2007. N Engl J Med 364:2016–2025. doi:10.1056/NEJMoa1005384. - DOI - PubMed
-
- Faralla C, Metruccio MM, De Chiara M, Mu R, Patras KA, Muzzi A, Grandi G, Margarit I, Doran KS, Janulczyk R. 2014. Analysis of two-component systems in group B Streptococcus shows that RgfAC and the novel FspSR modulate virulence and bacterial fitness. mBio 5:e00870-14. doi:10.1128/mBio.00870-14. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
