

MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health
- PMID: 29686086
- PMCID: PMC5984513
- DOI: 10.1073/pnas.1722335115
MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health
Abstract
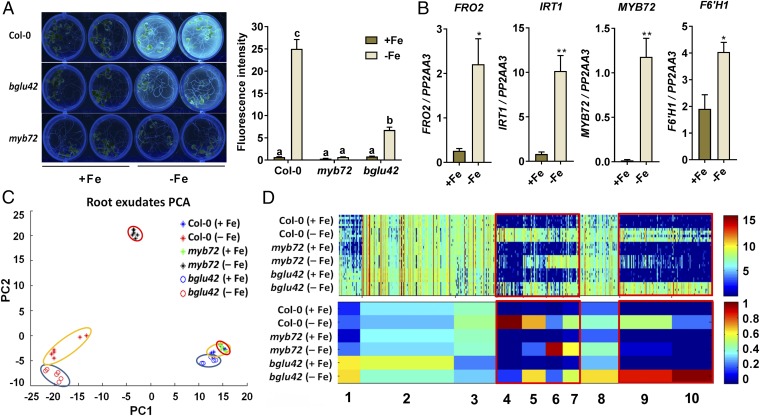
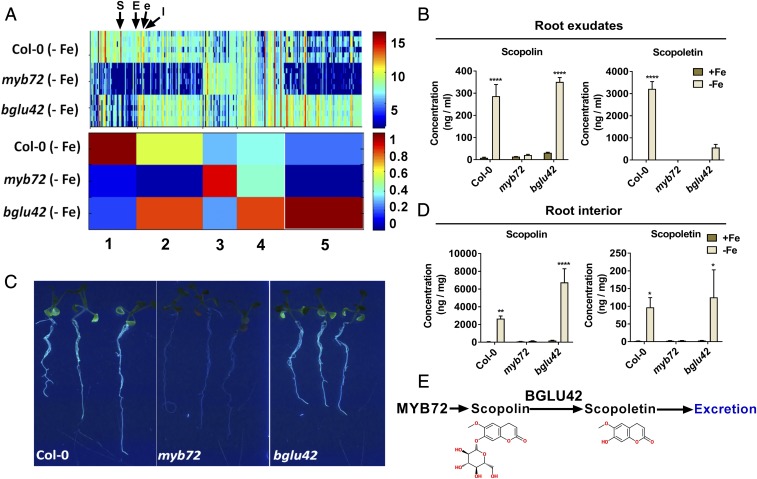
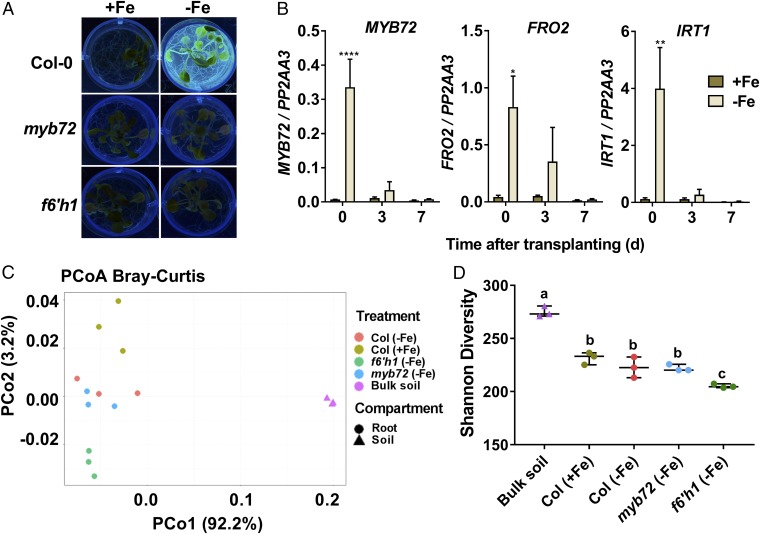
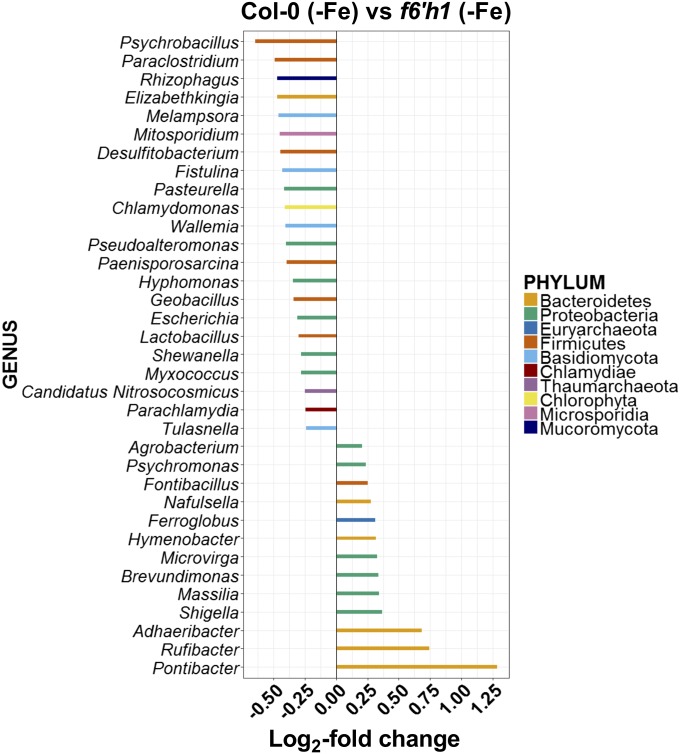
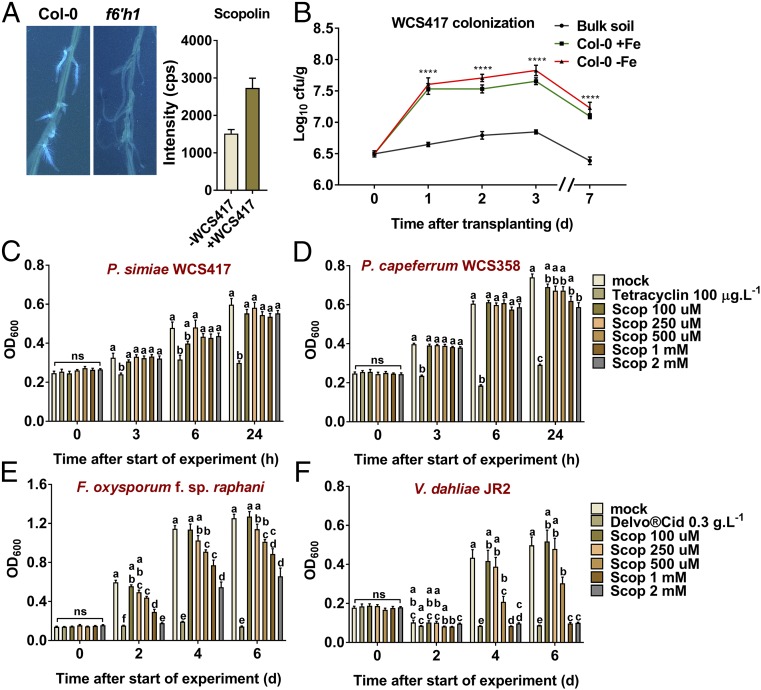
Plant roots nurture a tremendous diversity of microbes via exudation of photosynthetically fixed carbon sources. In turn, probiotic members of the root microbiome promote plant growth and protect the host plant against pathogens and pests. In the Arabidopsis thaliana-Pseudomonas simiae WCS417 model system the root-specific transcription factor MYB72 and the MYB72-controlled β-glucosidase BGLU42 emerged as important regulators of beneficial rhizobacteria-induced systemic resistance (ISR) and iron-uptake responses. MYB72 regulates the biosynthesis of iron-mobilizing fluorescent phenolic compounds, after which BGLU42 activity is required for their excretion into the rhizosphere. Metabolite fingerprinting revealed the antimicrobial coumarin scopoletin as a dominant metabolite that is produced in the roots and excreted into the rhizosphere in a MYB72- and BGLU42-dependent manner. Shotgun-metagenome sequencing of root-associated microbiota of Col-0, myb72, and the scopoletin biosynthesis mutant f6'h1 showed that scopoletin selectively impacts the assembly of the microbial community in the rhizosphere. We show that scopoletin selectively inhibits the soil-borne fungal pathogens Fusarium oxysporum and Verticillium dahliae, while the growth-promoting and ISR-inducing rhizobacteria P. simiae WCS417 and Pseudomonas capeferrum WCS358 are highly tolerant of the antimicrobial effect of scopoletin. Collectively, our results demonstrate a role for coumarins in microbiome assembly and point to a scenario in which plants and probiotic rhizobacteria join forces to trigger MYB72/BGLU42-dependent scopolin production and scopoletin excretion, resulting in improved niche establishment for the microbial partner and growth and immunity benefits for the host plant.
Keywords: coumarin; induced systemic resistance; iron-deficiency response; microbiome assembly; root metabolome.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Root-exuded coumarin shapes the root microbiome.Proc Natl Acad Sci U S A. 2018 May 29;115(22):5629-5631. doi: 10.1073/pnas.1805944115. Epub 2018 May 15. Proc Natl Acad Sci U S A. 2018. PMID: 29764997 Free PMC article. No abstract available.
References
-
- Bais HP, Weir TL, Perry LG, Gilroy S, Vivanco JM. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu Rev Plant Biol. 2006;57:233–266. - PubMed
-
- Berendsen RL, Pieterse CMJ, Bakker PAHM. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012;17:478–486. - PubMed
-
- Mendes R, et al. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science. 2011;332:1097–1100. - PubMed
-
- Bulgarelli D, Schlaeppi K, Spaepen S, Ver Loren van Themaat E, Schulze-Lefert P. Structure and functions of the bacterial microbiota of plants. Annu Rev Plant Biol. 2013;64:807–838. - PubMed
-
- Hacquard S, Spaepen S, Garrido-Oter R, Schulze-Lefert P. Interplay between innate immunity and the plant microbiota. Annu Rev Phytopathol. 2017;55:565–589. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
