

Hierarchical recruitment of ribosomal proteins and assembly factors remodels nucleolar pre-60S ribosomes
- PMID: 29691304
- PMCID: PMC6028539
- DOI: 10.1083/jcb.201711037
Hierarchical recruitment of ribosomal proteins and assembly factors remodels nucleolar pre-60S ribosomes
Abstract
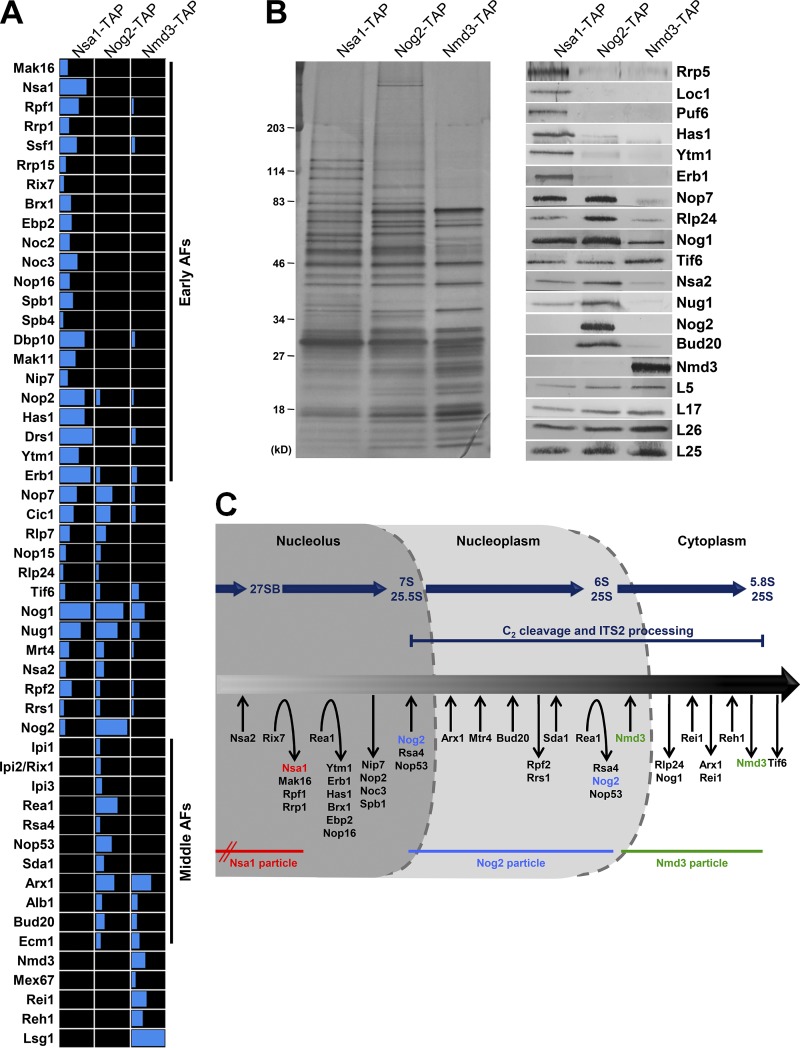
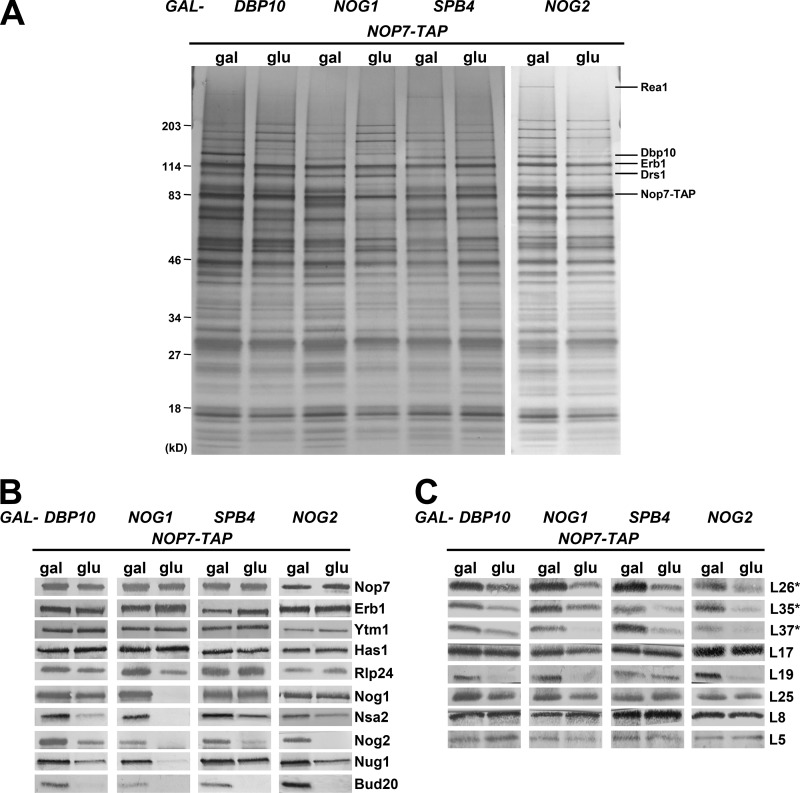
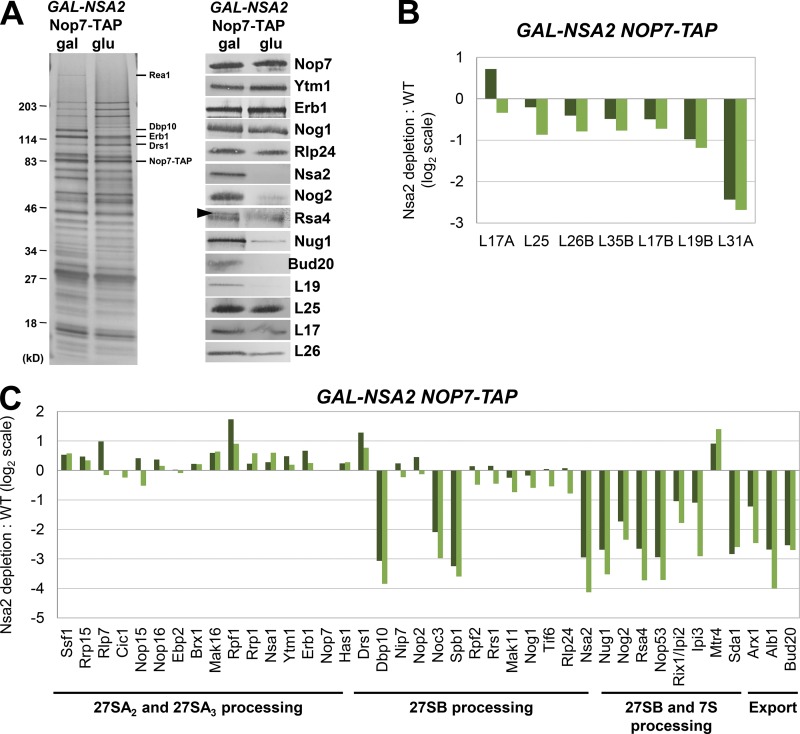
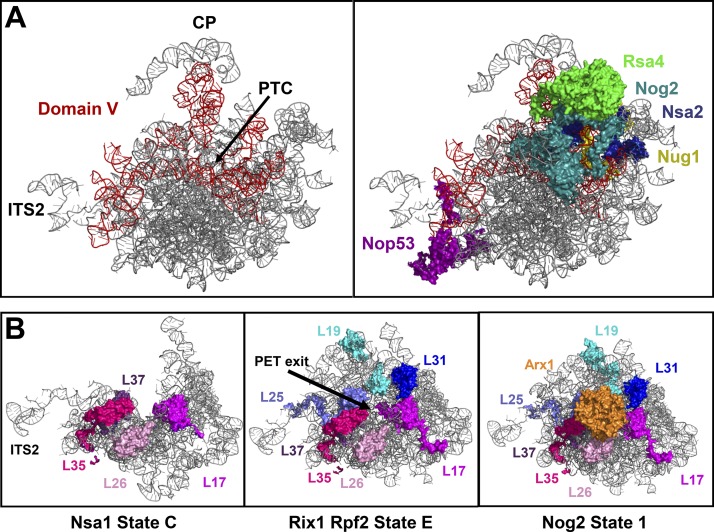
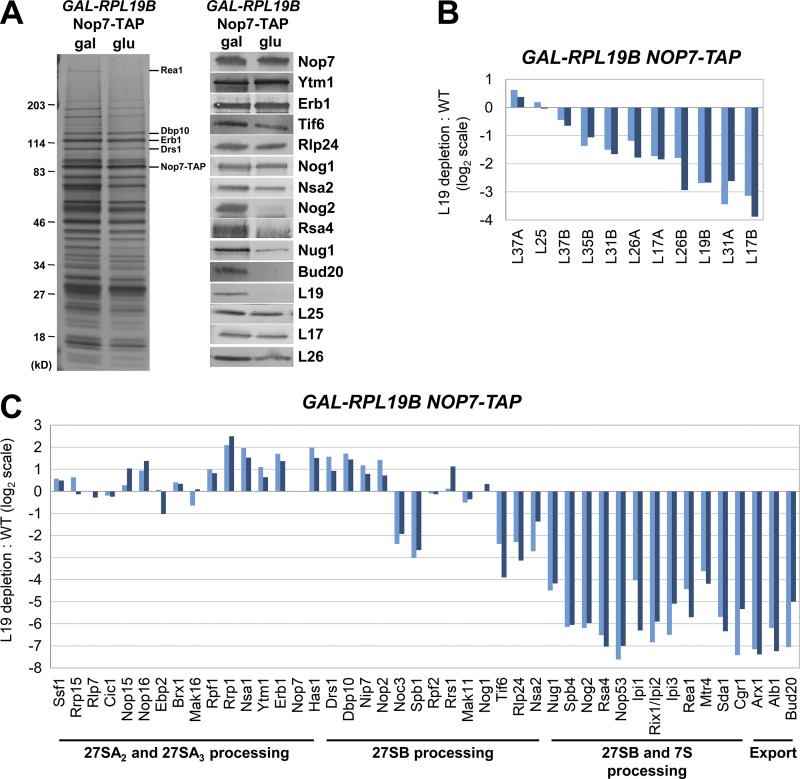
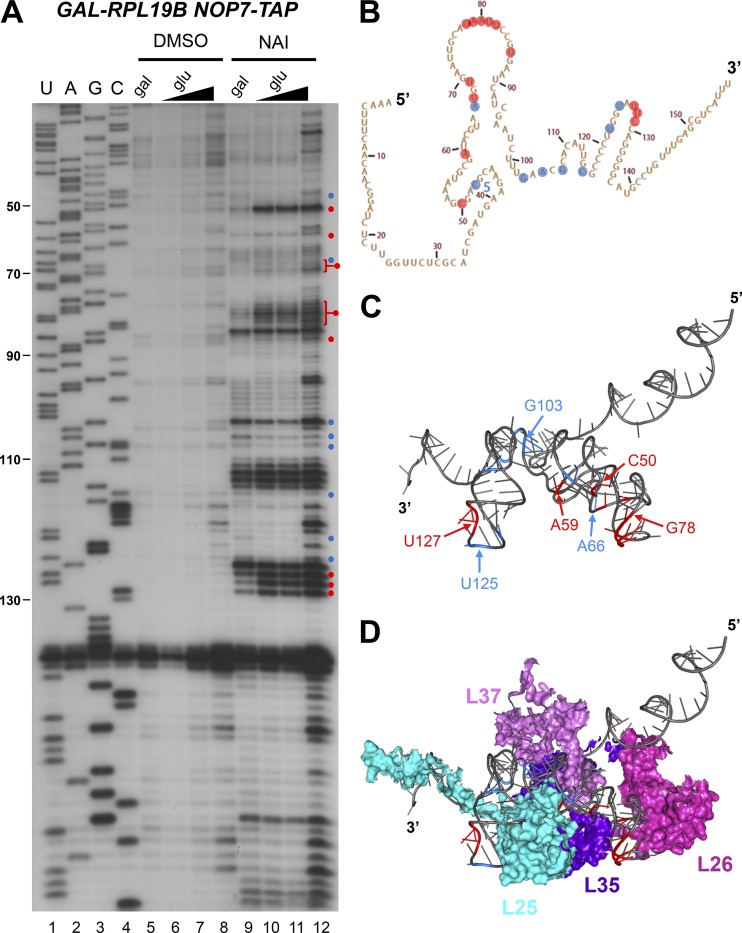
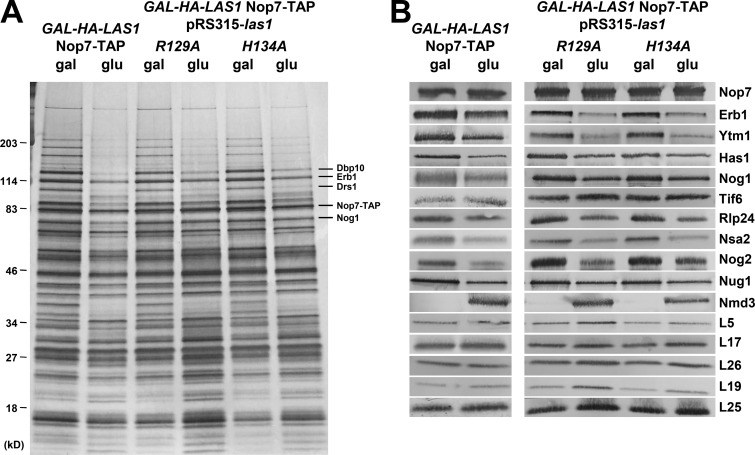
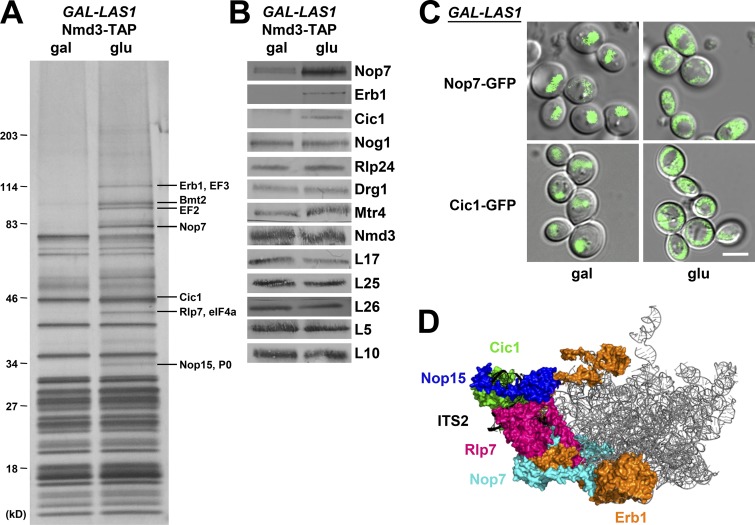
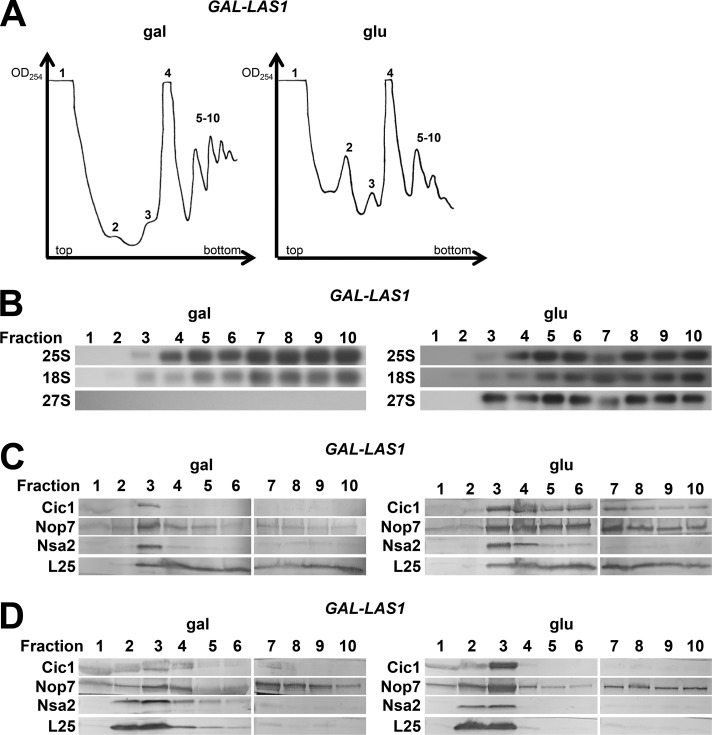
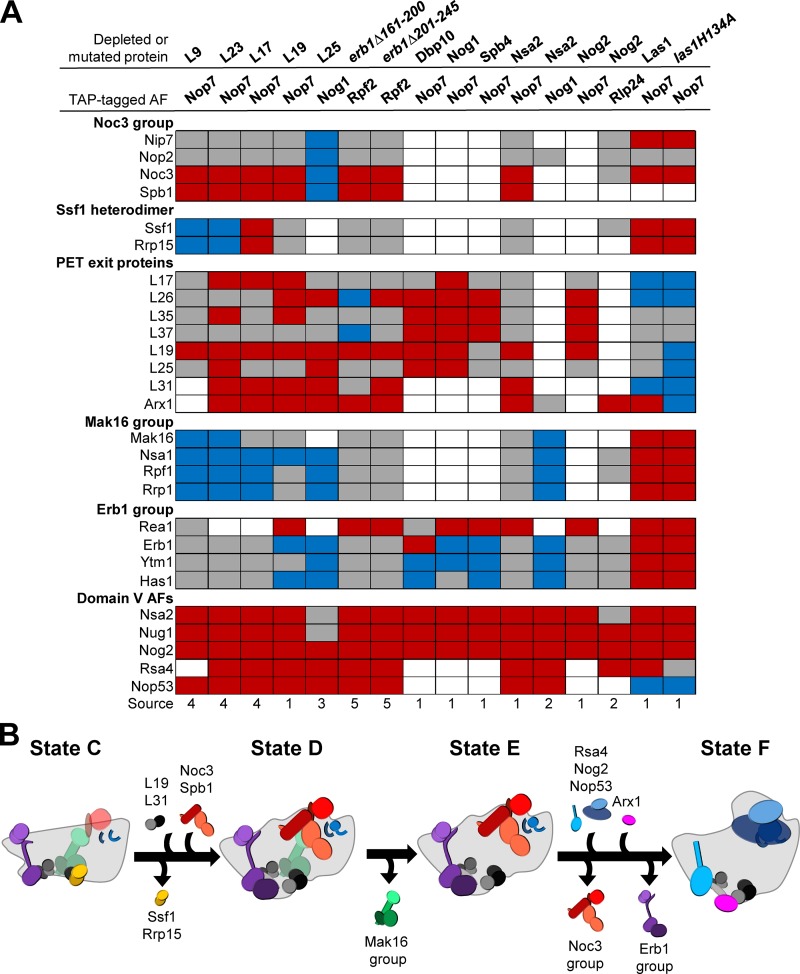
Ribosome biogenesis involves numerous preribosomal RNA (pre-rRNA) processing events to remove internal and external transcribed spacer sequences, ultimately yielding three mature rRNAs. Removal of the internal transcribed spacer 2 spacer RNA is the final step in large subunit pre-rRNA processing and begins with endonucleolytic cleavage at the C2 site of 27SB pre-rRNA. C2 cleavage requires the hierarchical recruitment of 11 ribosomal proteins and 14 ribosome assembly factors. However, the function of these proteins in C2 cleavage remained unclear. In this study, we have performed a detailed analysis of the effects of depleting proteins required for C2 cleavage and interpreted these results using cryo-electron microscopy structures of assembling 60S subunits. This work revealed that these proteins are required for remodeling of several neighborhoods, including two major functional centers of the 60S subunit, suggesting that these remodeling events form a checkpoint leading to C2 cleavage. Interestingly, when C2 cleavage is directly blocked by depleting or inactivating the C2 endonuclease, assembly progresses through all other subsequent steps.
© 2018 Biedka et al.
Figures
References
-
- Ausubel F.M., Brent R., Kingston R.E., Moore D.D., Seidman J.G., Smith J.A., and Struhl K.. 1994. Current protocols in molecular biology. John Wiley & Sons Inc, New York.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
