

Wnt Signaling Mediates LTP-Dependent Spine Plasticity and AMPAR Localization through Frizzled-7 Receptors
- PMID: 29694885
- PMCID: PMC5946458
- DOI: 10.1016/j.celrep.2018.03.119
Wnt Signaling Mediates LTP-Dependent Spine Plasticity and AMPAR Localization through Frizzled-7 Receptors
Abstract
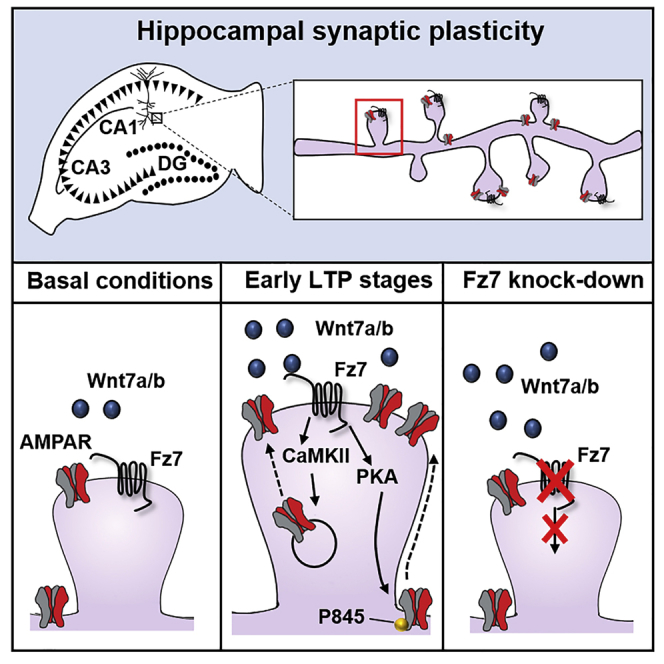
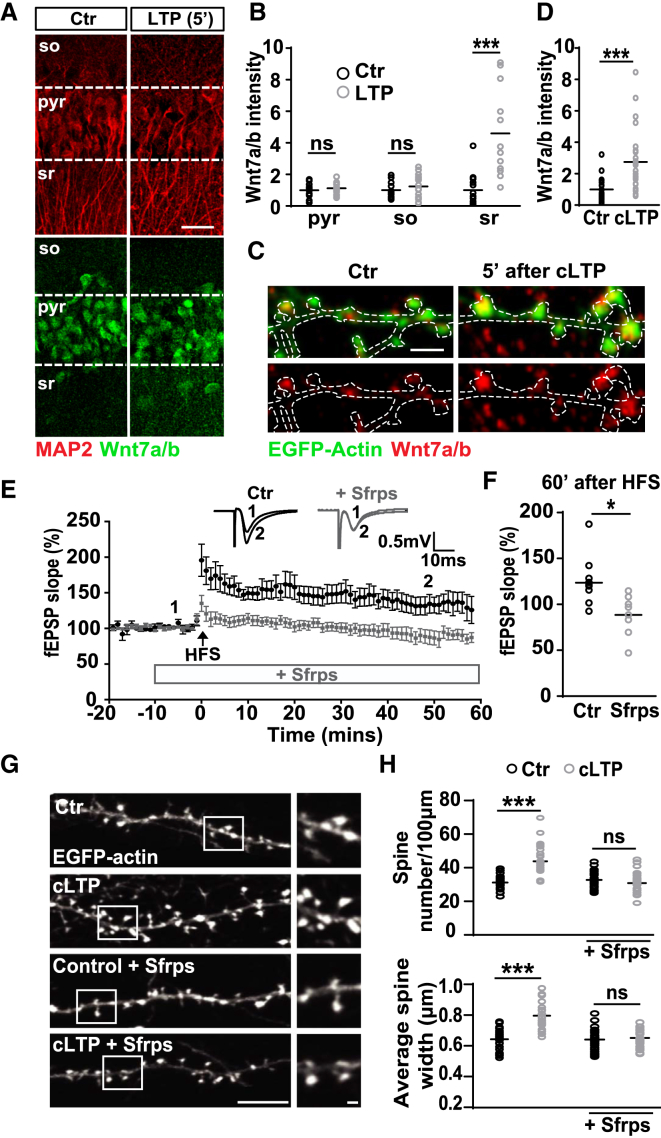
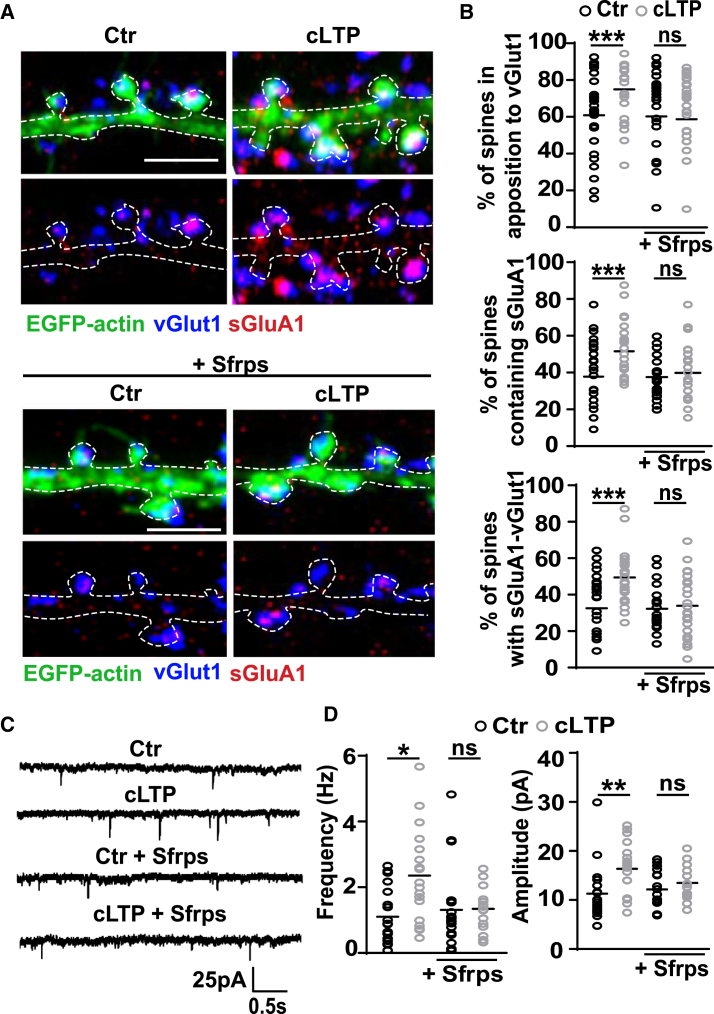
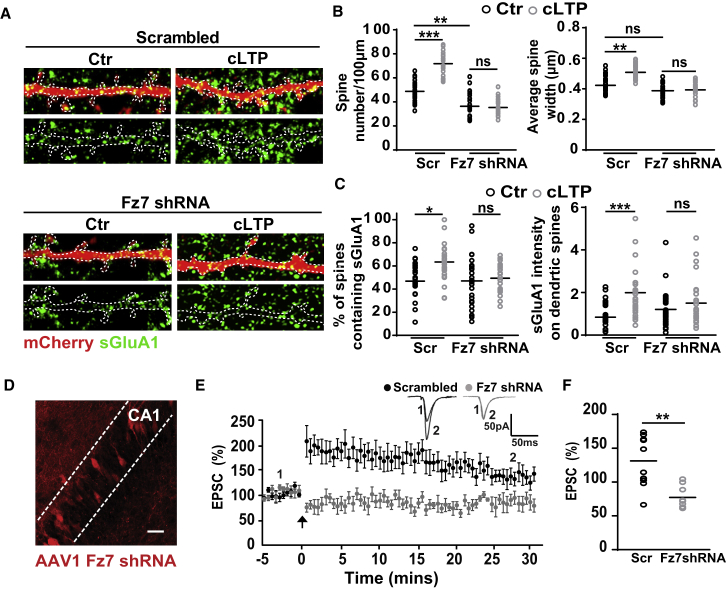
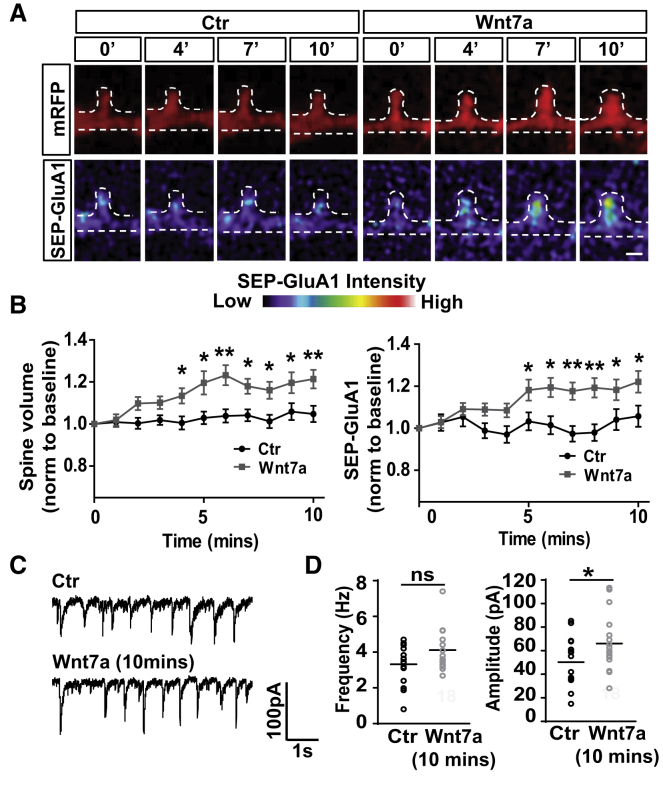
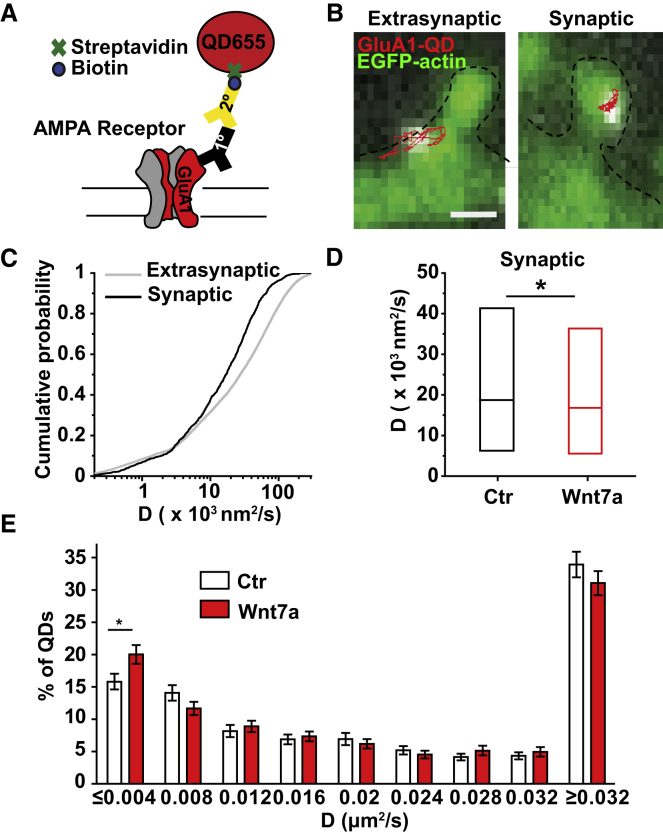
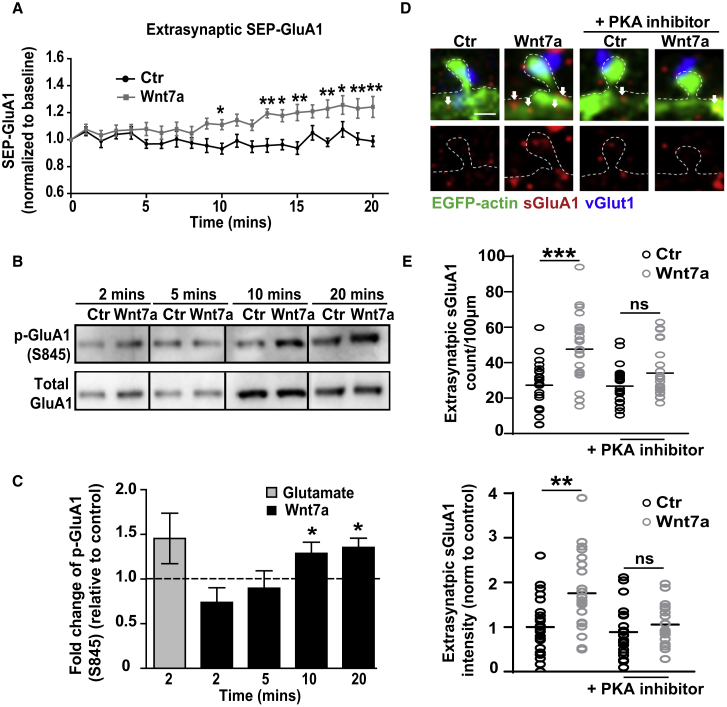
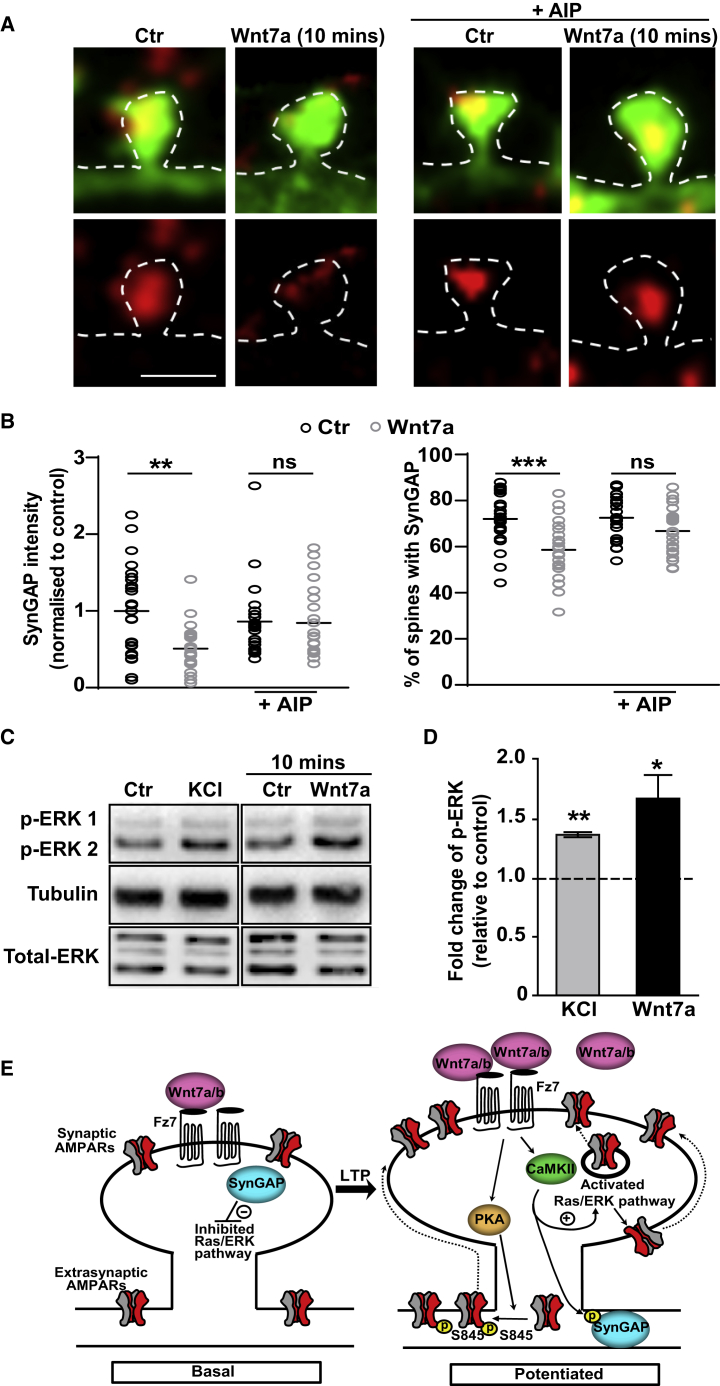
The structural and functional plasticity of synapses is critical for learning and memory. Long-term potentiation (LTP) induction promotes spine growth and AMPAR accumulation at excitatory synapses, leading to increased synaptic strength. Glutamate initiates these processes, but the contribution from extracellular modulators is not fully established. Wnts are required for spine formation; however, their impact on activity-mediated spine plasticity and AMPAR localization is unknown. We found that LTP induction rapidly increased synaptic Wnt7a/b protein levels. Acute blockade of endogenous Wnts or loss of postsynaptic Frizzled-7 (Fz7) receptors impaired LTP-mediated synaptic strength, spine growth, and AMPAR localization at synapses. Live imaging of SEP-GluA1 and single-particle tracking revealed that Wnt7a rapidly promoted synaptic AMPAR recruitment and trapping. Wnt7a, through Fz7, induced CaMKII-dependent loss of SynGAP from spines and increased extrasynaptic AMPARs by PKA phosphorylation. We identify a critical role for Wnt-Fz7 signaling in LTP-mediated synaptic accumulation of AMPARs and spine plasticity.
Keywords: AMPA receptors; Frizzled-7; LTP; Sfrps; Wnt signaling; spine plasticity; synaptic plasticity.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
