

An atypical N-ethylmaleimide sensitive factor enables the viability of nematode-resistant Rhg1 soybeans
- PMID: 29695628
- PMCID: PMC5948960
- DOI: 10.1073/pnas.1717070115
An atypical N-ethylmaleimide sensitive factor enables the viability of nematode-resistant Rhg1 soybeans
Abstract
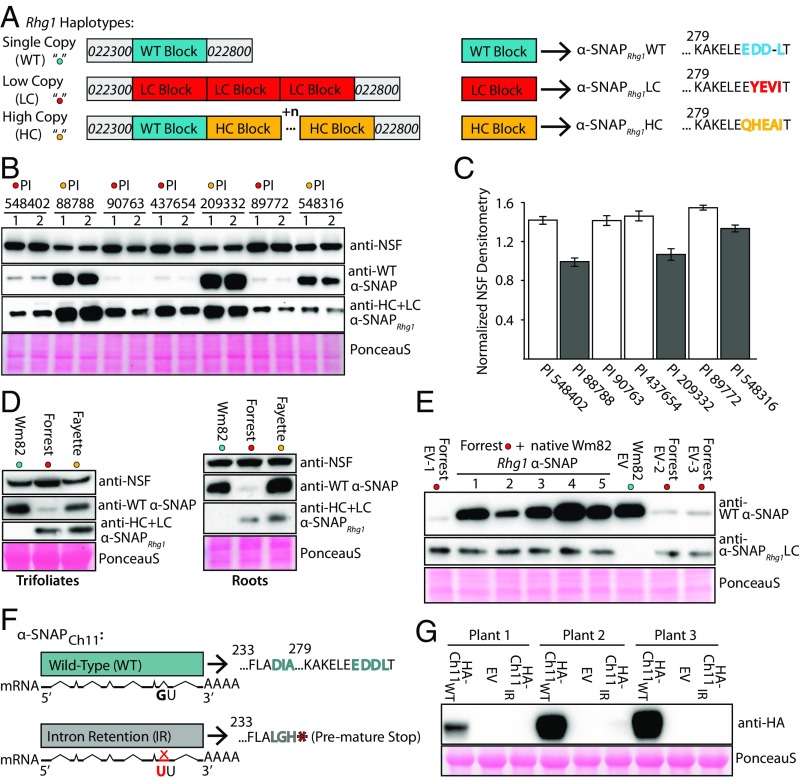
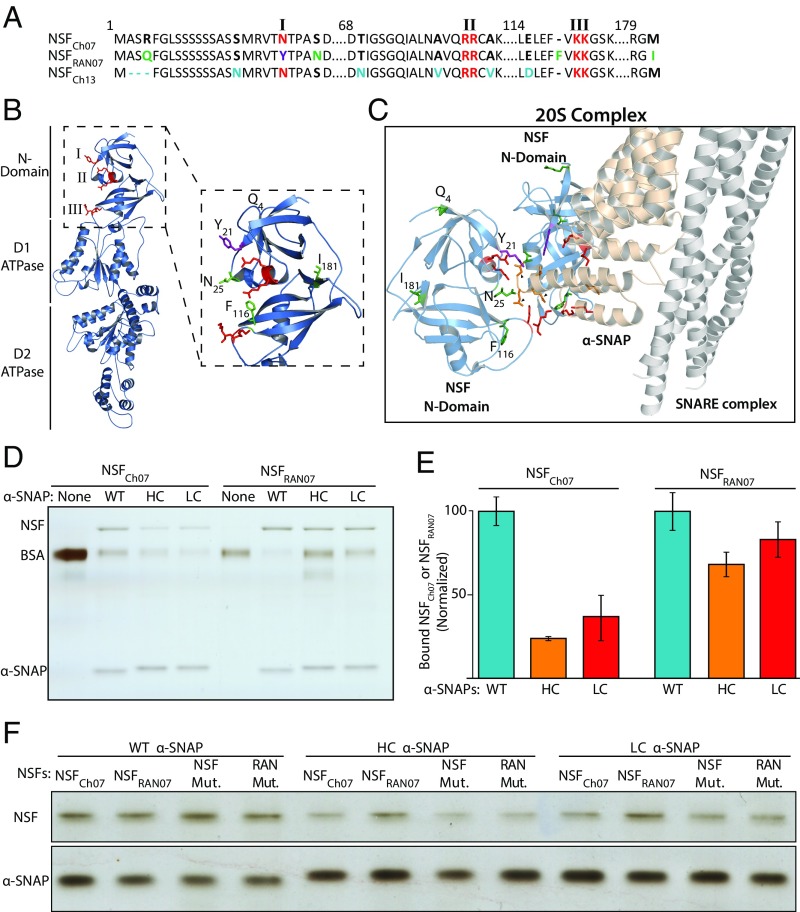
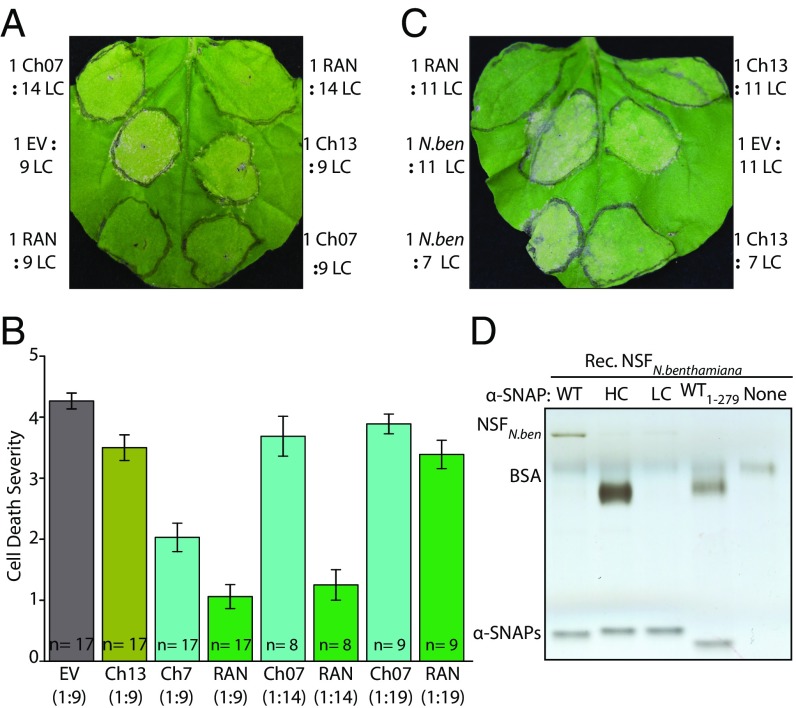
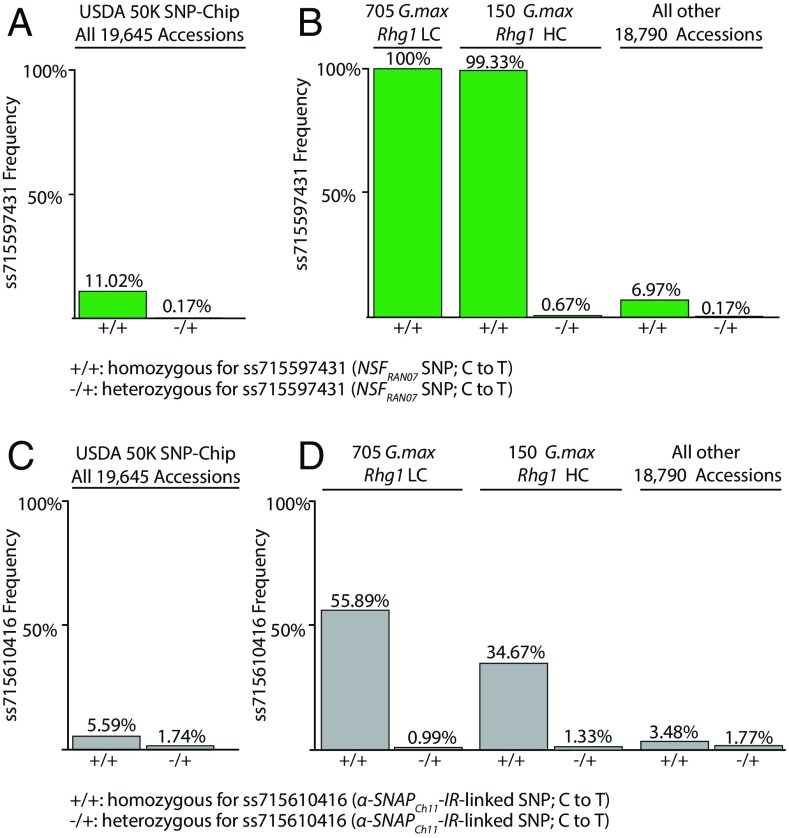
N-ethylmaleimide sensitive factor (NSF) and α-soluble NSF attachment protein (α-SNAP) are essential eukaryotic housekeeping proteins that cooperatively function to sustain vesicular trafficking. The "resistance to Heterodera glycines 1" (Rhg1) locus of soybean (Glycine max) confers resistance to soybean cyst nematode, a highly damaging soybean pest. Rhg1 loci encode repeat copies of atypical α-SNAP proteins that are defective in promoting NSF function and are cytotoxic in certain contexts. Here, we discovered an unusual NSF allele (Rhg1-associated NSF on chromosome 07; NSFRAN07 ) in Rhg1+ germplasm. NSFRAN07 protein modeling to mammalian NSF/α-SNAP complex structures indicated that at least three of the five NSFRAN07 polymorphisms reside adjacent to the α-SNAP binding interface. NSFRAN07 exhibited stronger in vitro binding with Rhg1 resistance-type α-SNAPs. NSFRAN07 coexpression in planta was more protective against Rhg1 α-SNAP cytotoxicity, relative to WT NSFCh07 Investigation of a previously reported segregation distortion between chromosome 18 Rhg1 and a chromosome 07 interval now known to contain the Glyma.07G195900 NSF gene revealed 100% coinheritance of the NSFRAN07 allele with disease resistance Rhg1 alleles, across 855 soybean accessions and in all examined Rhg1+ progeny from biparental crosses. Additionally, we show that some Rhg1-mediated resistance is associated with depletion of WT α-SNAP abundance via selective loss of WT α-SNAP loci. Hence atypical coevolution of the soybean SNARE-recycling machinery has balanced the acquisition of an otherwise disruptive housekeeping protein, enabling a valuable disease resistance trait. Our findings further indicate that successful engineering of Rhg1-related resistance in plants will require a compatible NSF partner for the resistance-conferring α-SNAP.
Keywords: NSF; Rhg1; plant disease resistance; soybean cyst nematode; α-SNAP.
Conflict of interest statement
Conflict of interest statement: A patent application covering the presently described work has been filed by the Wisconsin Alumni Research Foundation.
Figures
References
-
- Kyndt T, Vieira P, Gheysen G, de Almeida-Engler J. Nematode feeding sites: Unique organs in plant roots. Planta. 2013;238:807–818. - PubMed
-
- Niblack TL, Lambert KN, Tylka GL. A model plant pathogen from the kingdom Animalia: Heterodera glycines, the soybean cyst nematode. Annu Rev Phytopathol. 2006;44:283–303. - PubMed
-
- Mitchum MG. Soybean resistance to the soybean cyst nematode Heterodera glycines: An update. Phytopathology. 2016;106:1444–1450. - PubMed
-
- Allen TW, et al. Soybean yield loss estimates due to diseases in the United States and Ontario, Canada, from 2010 to 2014. Plant Health Prog. 2017;18:19–27.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
