

The multiple myeloma risk allele at 5q15 lowers ELL2 expression and increases ribosomal gene expression
- PMID: 29695719
- PMCID: PMC5917026
- DOI: 10.1038/s41467-018-04082-2
The multiple myeloma risk allele at 5q15 lowers ELL2 expression and increases ribosomal gene expression
Abstract
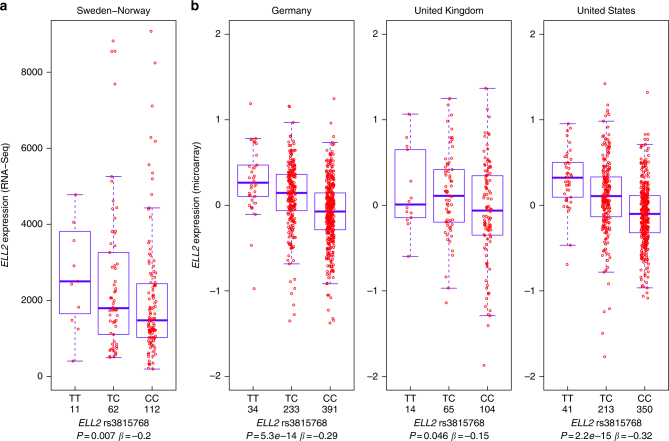
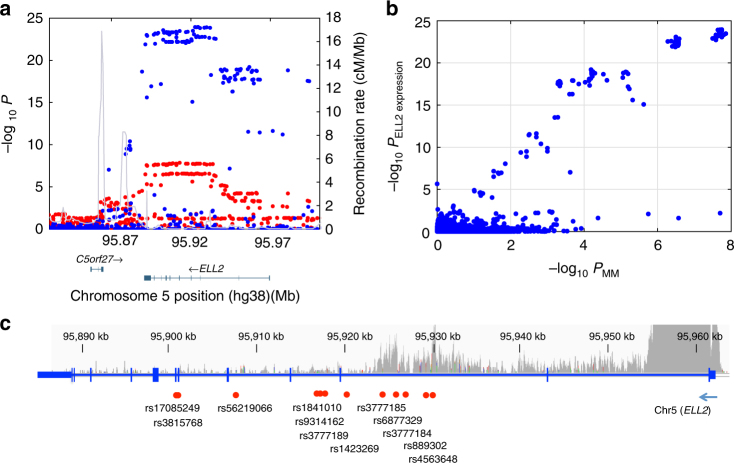
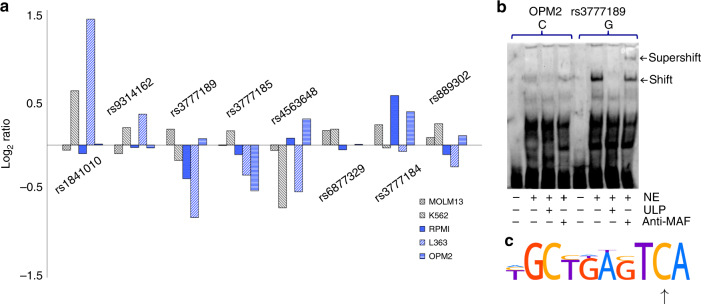
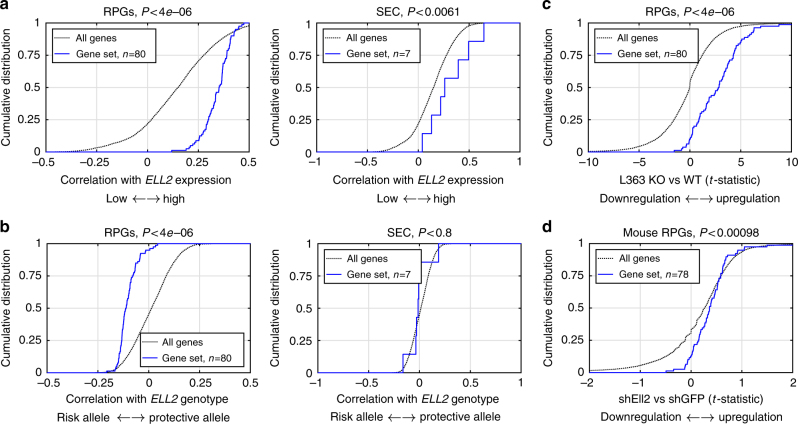
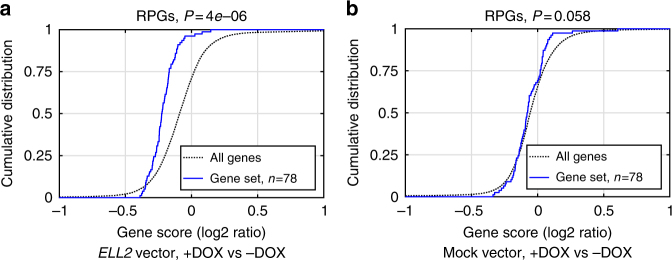
Recently, we identified ELL2 as a susceptibility gene for multiple myeloma (MM). To understand its mechanism of action, we performed expression quantitative trait locus analysis in CD138+ plasma cells from 1630 MM patients from four populations. We show that the MM risk allele lowers ELL2 expression in these cells (Pcombined = 2.5 × 10-27; βcombined = -0.24 SD), but not in peripheral blood or other tissues. Consistent with this, several variants representing the MM risk allele map to regulatory genomic regions, and three yield reduced transcriptional activity in plasmocytoma cell lines. One of these (rs3777189-C) co-locates with the best-supported lead variants for ELL2 expression and MM risk, and reduces binding of MAFF/G/K family transcription factors. Moreover, further analysis reveals that the MM risk allele associates with upregulation of gene sets related to ribosome biogenesis, and knockout/knockdown and rescue experiments in plasmocytoma cell lines support a cause-effect relationship. Our results provide mechanistic insight into MM predisposition.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Genetic Predisposition to Multiple Myeloma at 5q15 Is Mediated by an ELL2 Enhancer Polymorphism.Cell Rep. 2017 Sep 12;20(11):2556-2564. doi: 10.1016/j.celrep.2017.08.062. Cell Rep. 2017. PMID: 28903037 Free PMC article.
-
Variants in ELL2 influencing immunoglobulin levels associate with multiple myeloma.Nat Commun. 2015 May 26;6:7213. doi: 10.1038/ncomms8213. Nat Commun. 2015. PMID: 26007630 Free PMC article.
-
Integrative Genomic Analysis Predicts Causative Cis-Regulatory Mechanisms of the Breast Cancer-Associated Genetic Variant rs4415084.Cancer Res. 2018 Apr 1;78(7):1579-1591. doi: 10.1158/0008-5472.CAN-17-3486. Epub 2018 Jan 19. Cancer Res. 2018. PMID: 29351903 Free PMC article.
-
Insights from the gene expression profiling of multiple myeloma.Curr Hematol Rep. 2004 Jan;3(1):67-73. Curr Hematol Rep. 2004. PMID: 14695854 Review.
-
Diagnosis and risk stratification in multiple myeloma.Hematol Oncol Clin North Am. 2014 Oct;28(5):791-813. doi: 10.1016/j.hoc.2014.06.007. Epub 2014 Jul 22. Hematol Oncol Clin North Am. 2014. PMID: 25212883 Review.
Cited by
-
ELL2 Influences Transcription Elongation, Splicing, Ig Secretion and Growth.J Mucosal Immunol Res. 2019;3(1):112. Epub 2019 Jul 12. J Mucosal Immunol Res. 2019. PMID: 31930204 Free PMC article.
-
Transcriptome Analysis of Diffuse Large B-Cell Lymphoma Cells Inducibly Expressing MyD88 L265P Mutation Identifies Upregulated CD44, LGALS3, NFKBIZ, and BATF as Downstream Targets of Oncogenic NF-κB Signaling.Int J Mol Sci. 2023 Mar 15;24(6):5623. doi: 10.3390/ijms24065623. Int J Mol Sci. 2023. PMID: 36982699 Free PMC article.
-
A meta-analysis of genome-wide association studies of multiple myeloma among men and women of African ancestry.Blood Adv. 2020 Jan 14;4(1):181-190. doi: 10.1182/bloodadvances.2019000491. Blood Adv. 2020. PMID: 31935283 Free PMC article.
-
Multiple Myeloma: Bioinformatic Analysis for Identification of Key Genes and Pathways.Bioinform Biol Insights. 2022 Aug 6;16:11779322221115545. doi: 10.1177/11779322221115545. eCollection 2022. Bioinform Biol Insights. 2022. PMID: 35958298 Free PMC article.
-
Germline variants at SOHLH2 influence multiple myeloma risk.Blood Cancer J. 2021 Apr 19;11(4):76. doi: 10.1038/s41408-021-00468-6. Blood Cancer J. 2021. PMID: 33875642 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
