

Dnmt2 mediates intergenerational transmission of paternally acquired metabolic disorders through sperm small non-coding RNAs
- PMID: 29695786
- PMCID: PMC5926820
- DOI: 10.1038/s41556-018-0087-2
Dnmt2 mediates intergenerational transmission of paternally acquired metabolic disorders through sperm small non-coding RNAs
Abstract
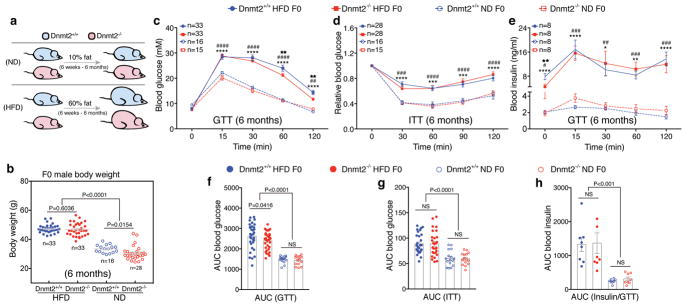
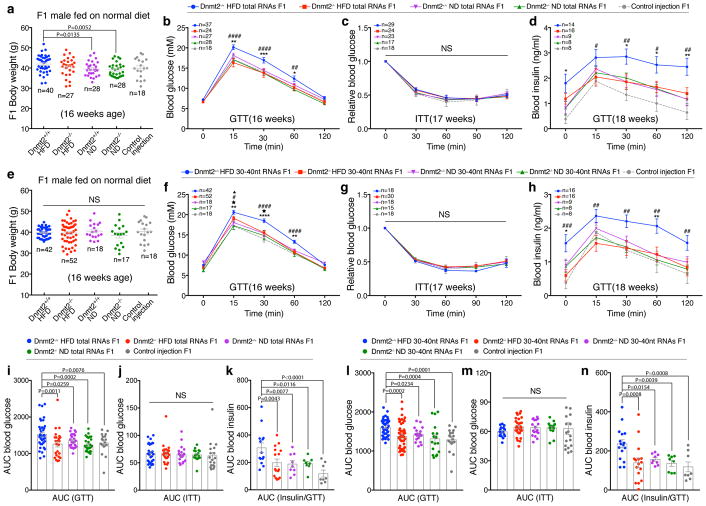
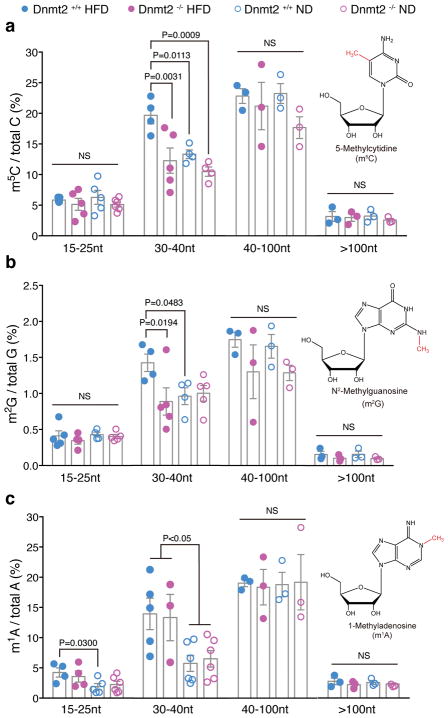
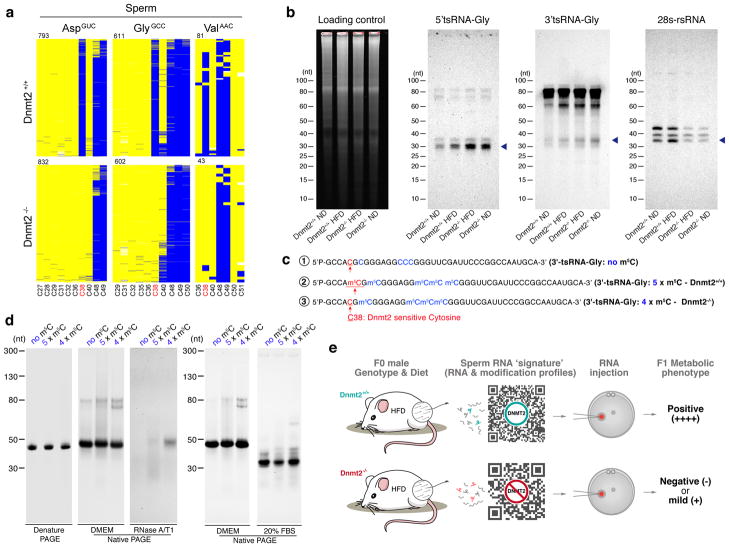
The discovery of RNAs (for example, messenger RNAs, non-coding RNAs) in sperm has opened the possibility that sperm may function by delivering additional paternal information aside from solely providing the DNA 1 . Increasing evidence now suggests that sperm small non-coding RNAs (sncRNAs) can mediate intergenerational transmission of paternally acquired phenotypes, including mental stress2,3 and metabolic disorders4-6. How sperm sncRNAs encode paternal information remains unclear, but the mechanism may involve RNA modifications. Here we show that deletion of a mouse tRNA methyltransferase, DNMT2, abolished sperm sncRNA-mediated transmission of high-fat-diet-induced metabolic disorders to offspring. Dnmt2 deletion prevented the elevation of RNA modifications (m5C, m2G) in sperm 30-40 nt RNA fractions that are induced by a high-fat diet. Also, Dnmt2 deletion altered the sperm small RNA expression profile, including levels of tRNA-derived small RNAs and rRNA-derived small RNAs, which might be essential in composing a sperm RNA 'coding signature' that is needed for paternal epigenetic memory. Finally, we show that Dnmt2-mediated m5C contributes to the secondary structure and biological properties of sncRNAs, implicating sperm RNA modifications as an additional layer of paternal hereditary information.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Dad's diet - smRNA methylation signatures in sperm pass on disease risk.Nat Rev Endocrinol. 2018 Aug;14(8):446-447. doi: 10.1038/s41574-018-0051-z. Nat Rev Endocrinol. 2018. PMID: 29959429 No abstract available.
References
-
- Chen Q, et al. Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder. Science. 2016;351:397–400. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
