

Trypanosomal mitochondrial intermediate peptidase does not behave as a classical mitochondrial processing peptidase
- PMID: 29698456
- PMCID: PMC5919513
- DOI: 10.1371/journal.pone.0196474
Trypanosomal mitochondrial intermediate peptidase does not behave as a classical mitochondrial processing peptidase
Abstract
Upon their translocation into the mitochondrial matrix, the N-terminal pre-sequence of nuclear-encoded proteins undergoes cleavage by mitochondrial processing peptidases. Some proteins require more than a single processing step, which involves several peptidases. Down-regulation of the putative Trypanosoma brucei mitochondrial intermediate peptidase (MIP) homolog by RNAi renders the cells unable to grow after 48 hours of induction. Ablation of MIP results in the accumulation of the precursor of the trypanosomatid-specific trCOIV protein, the largest nuclear-encoded subunit of the cytochrome c oxidase complex in this flagellate. However, the trCOIV precursor of the same size accumulates also in trypanosomes in which either alpha or beta subunits of the mitochondrial processing peptidase (MPP) have been depleted. Using a chimeric protein that consists of the N-terminal sequence of a putative subunit of respiratory complex I fused to a yellow fluorescent protein, we assessed the accumulation of the precursor protein in trypanosomes, in which RNAi was induced against the alpha or beta subunits of MPP or MIP. The observed accumulation of precursors indicates MIP depletion affects the activity of the cannonical MPP, or at least one of its subunits.
Conflict of interest statement
Figures
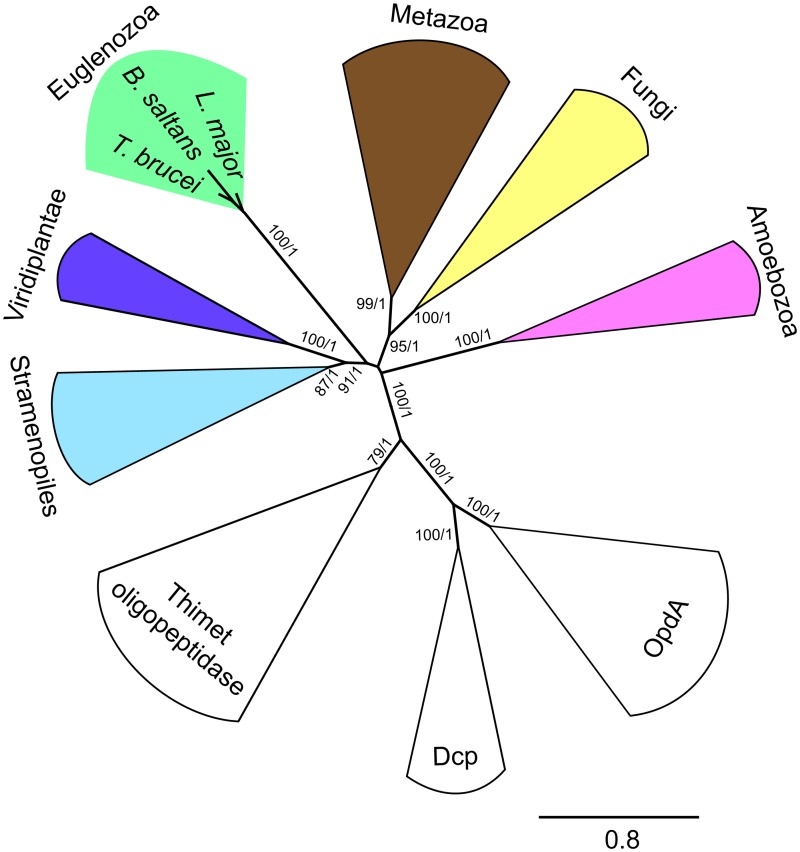
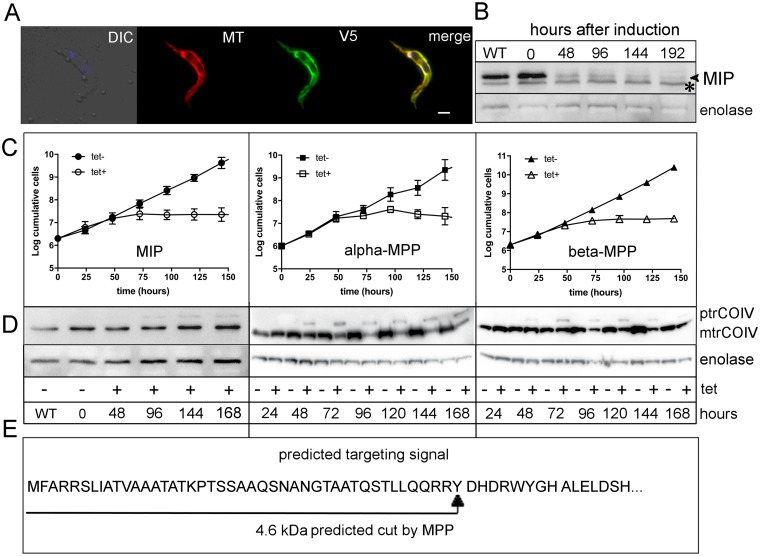
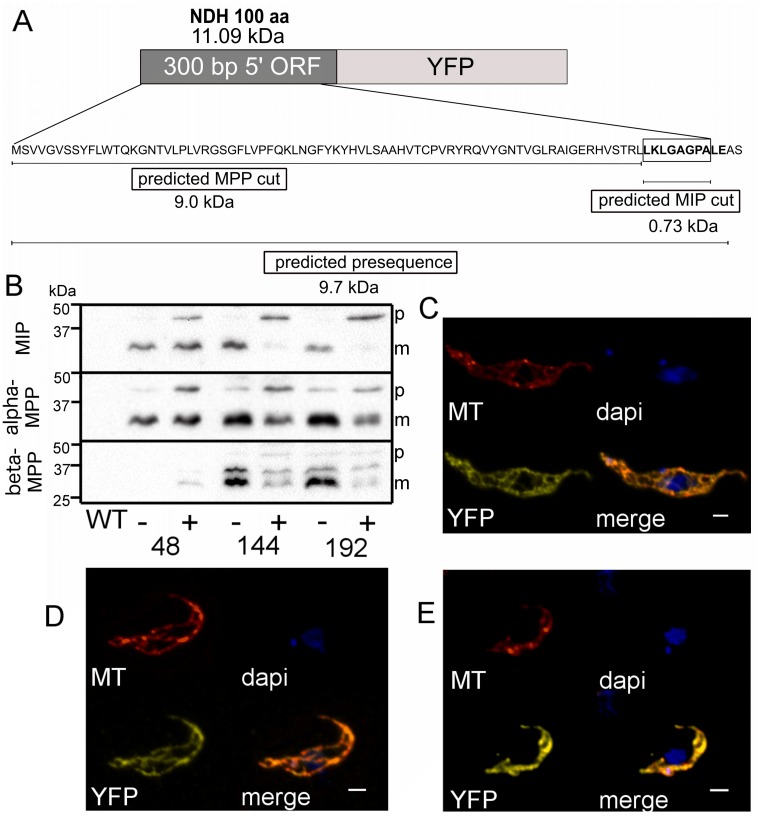
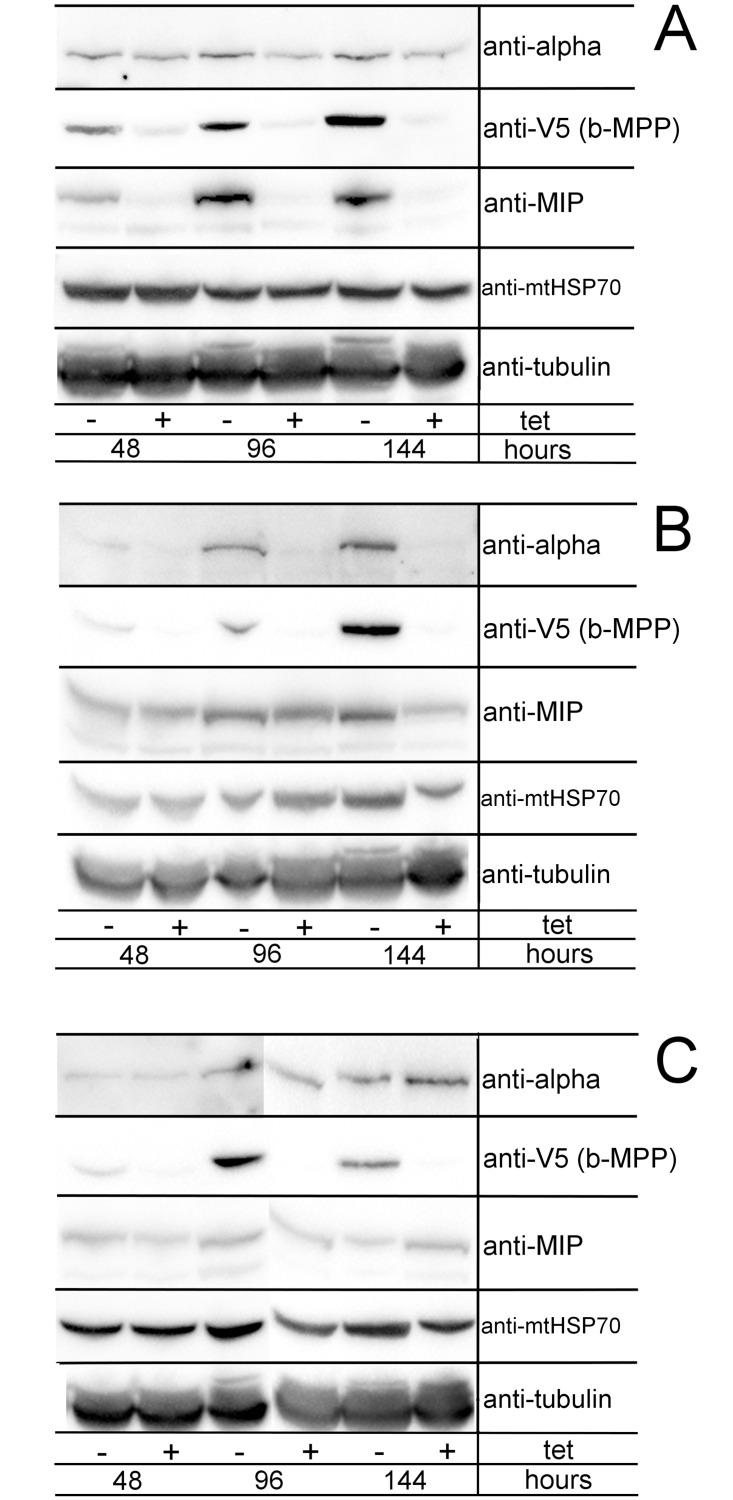
References
-
- Neupert W. Protein import into mitochondria. Annu Rev Biochem. 1997;66: 863–917. doi: 10.1146/annurev.biochem.66.1.863 - DOI - PubMed
-
- Schmidt O, Pfanner N, Meisinger C. Mitochondrial protein import: from proteomics to functional mechanisms. Nat Rev Mol Cell Biol. 2010;11: 655–667. doi: 10.1038/nrm2959 - DOI - PubMed
-
- Hildenbeutel M, Habib SJ, Herrmann JM, Rapaport D. New insights into the mechanism of precursor protein insertion into the mitochondrial membranes. Int Rev Cell Mol Biol. 2008;268: 147–190. doi: 10.1016/S1937-6448(08)00805-8 - DOI - PubMed
-
- Eckers E, Cyrklaff M, Simpson L, Deponte M. Mitochondrial protein import pathways are functionally conserved among eukaryotes despite compositional diversity of the import machineries. Biol Chem. 2012;393: 513–524. doi: 10.1515/hsz-2011-0255 - DOI - PubMed
-
- Chacinska A, Koehler CM, Milenkovic D, Lithgow T, Pfanner N. Importing mitochondrial proteins: machineries and mechanisms. Cell. 2009;138: 628–644. doi: 10.1016/j.cell.2009.08.005 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
