

Brain region-specific disruption of Shank3 in mice reveals a dissociation for cortical and striatal circuits in autism-related behaviors
- PMID: 29700290
- PMCID: PMC5919902
- DOI: 10.1038/s41398-018-0142-6
Brain region-specific disruption of Shank3 in mice reveals a dissociation for cortical and striatal circuits in autism-related behaviors
Abstract
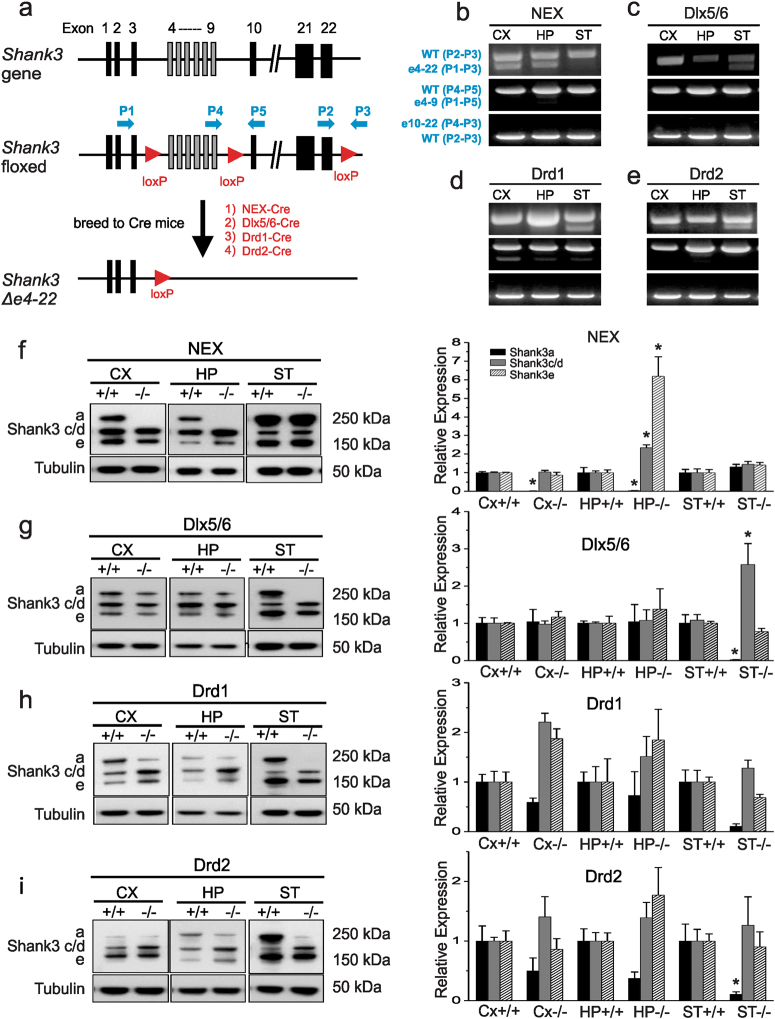
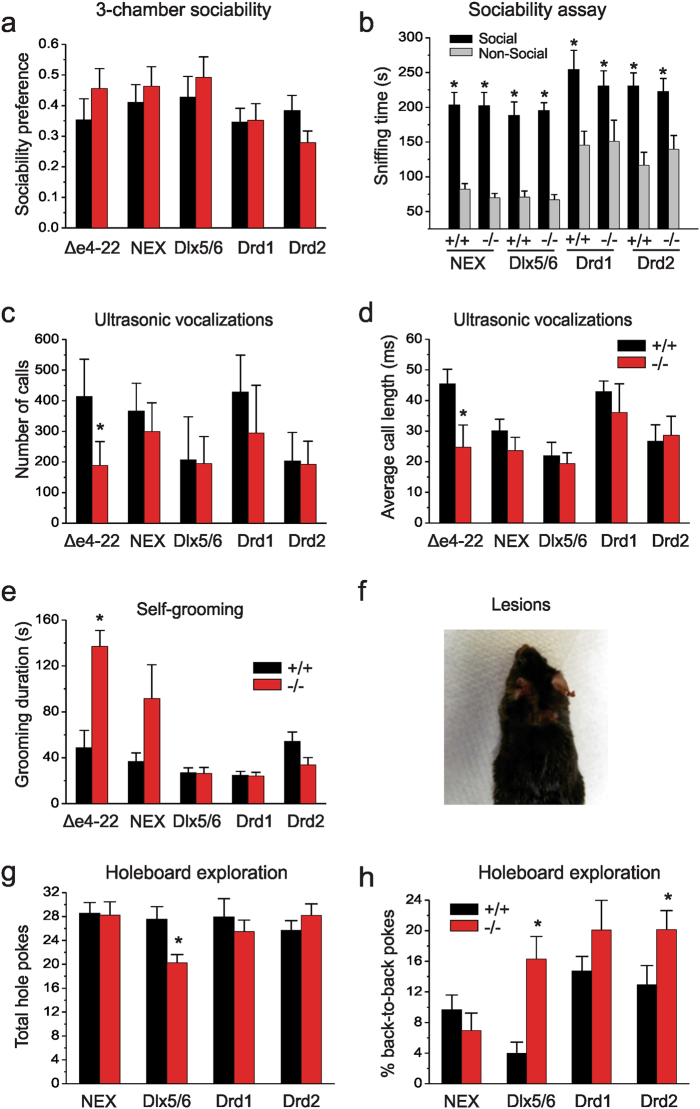
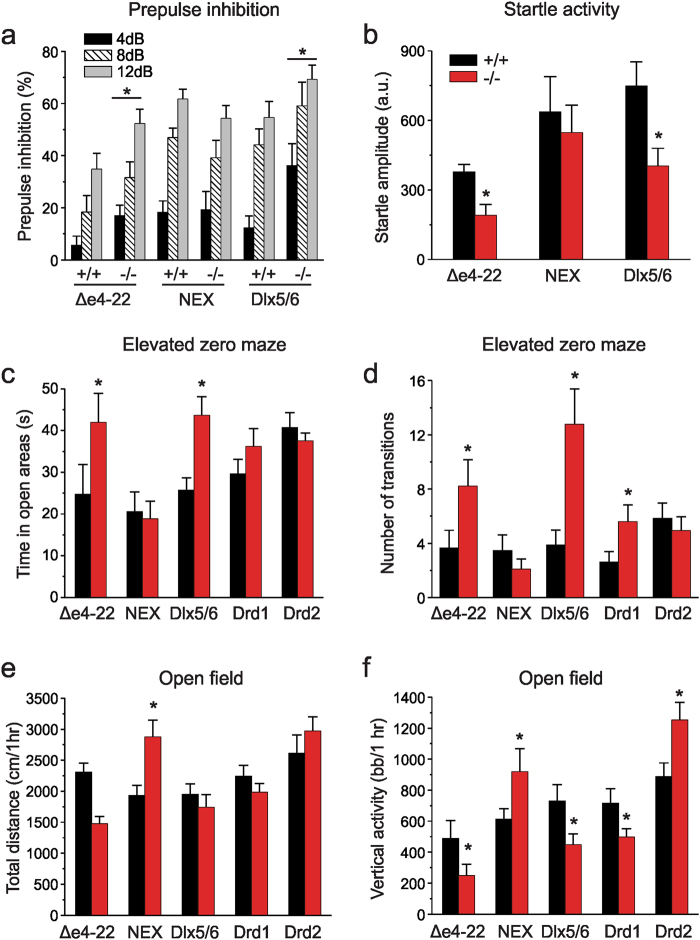
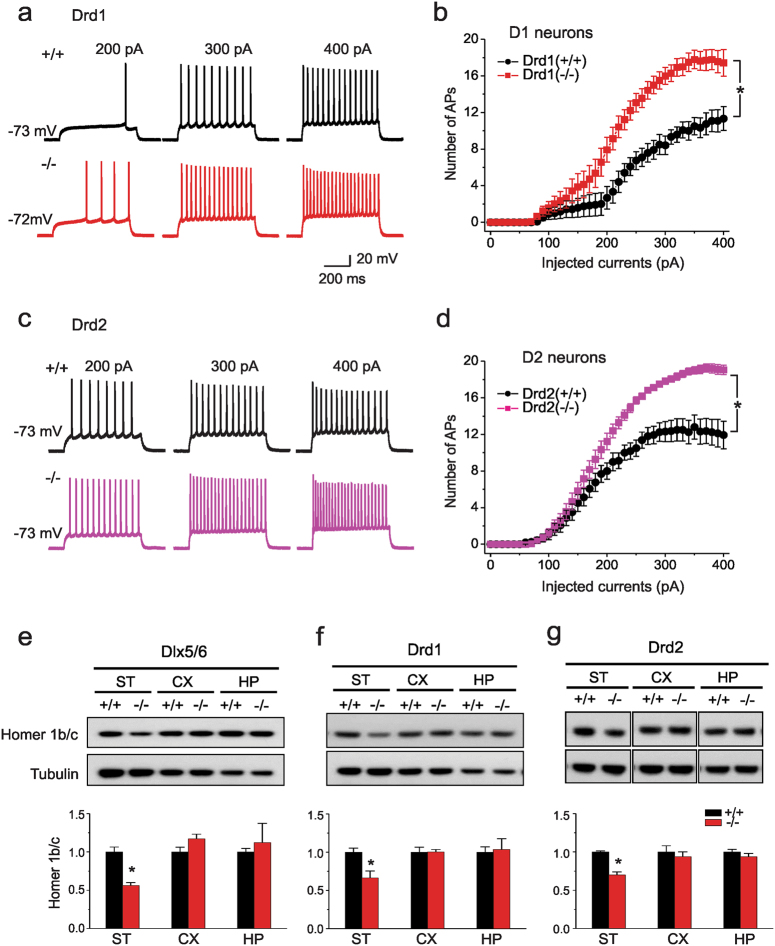
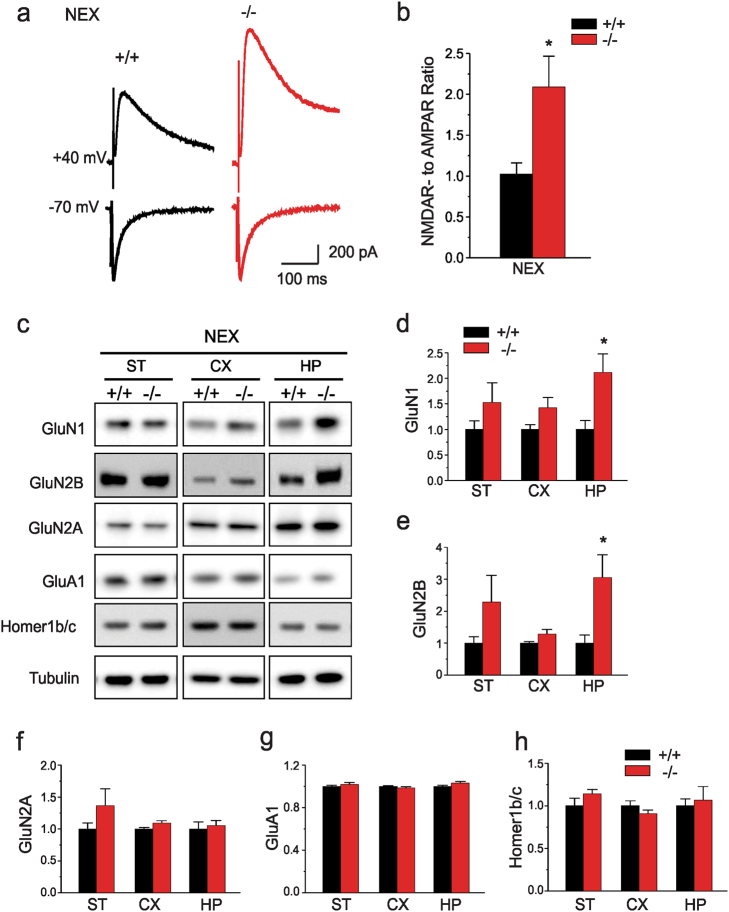
We previously reported a new line of Shank3 mutant mice which led to a complete loss of Shank3 by deleting exons 4-22 (Δe4-22) globally. Δe4-22 mice display robust ASD-like behaviors including impaired social interaction and communication, increased stereotypical behavior and excessive grooming, and a profound deficit in instrumental learning. However, the anatomical and neural circuitry underlying these behaviors are unknown. We generated mice with Shank3 selectively deleted in forebrain, striatum, and striatal D1 and D2 cells. These mice were used to interrogate the circuit/brain-region and cell-type specific role of Shank3 in the expression of autism-related behaviors. Whole-cell patch recording and biochemical analyses were used to study the synaptic function and molecular changes in specific brain regions. We found perseverative exploratory behaviors in mice with deletion of Shank3 in striatal inhibitory neurons. Conversely, self-grooming induced lesions were observed in mice with deletion of Shank3 in excitatory neurons of forebrain. However, social, communicative, and instrumental learning behaviors were largely unaffected in these mice, unlike what is seen in global Δe4-22 mice. We discovered unique patterns of change for the biochemical and electrophysiological findings in respective brain regions that reflect the complex nature of transcriptional regulation of Shank3. Reductions in Homer1b/c and membrane hyper-excitability were observed in striatal loss of Shank3. By comparison, Shank3 deletion in hippocampal neurons resulted in increased NMDAR-currents and GluN2B-containing NMDARs. These results together suggest that Shank3 may differentially regulate neural circuits that control behavior. Our study supports a dissociation of Shank3 functions in cortical and striatal neurons in ASD-related behaviors, and it illustrates the complexity of neural circuit mechanisms underlying these behaviors.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- American Psychiatric Association. Diagnostic and statistical manual of mental disorders. 5th edn. Arlington, VA: American Psychiatric Publishing; 2013.
-
- Autism Developmental Disabilities Monitoring Network. Prevalence of autism spectrum disorders--Autism and Developmental Disabilities Monitoring Network, 14 sites, United States, 2008. MMWR Surveill. Summ. 2012;61:1–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
