

The role of catecholamines in HIV neuropathogenesis
- PMID: 29705605
- PMCID: PMC6204123
- DOI: 10.1016/j.brainres.2018.04.030
The role of catecholamines in HIV neuropathogenesis
Abstract
The success of anti-retroviral therapy has improved the quality of life and lifespan of HIV + individuals, transforming HIV infection into a chronic condition. These improvements have come with a cost, as chronic HIV infection and long-term therapy have resulted in the emergence of a number of new pathologies. This includes a variety of the neuropathological and neurocognitive effects collectively known as HIVassociated neurocognitive disorders (HAND) or NeuroHIV. These effects persist even in the absence of viral replication, suggesting that they are mediated the long-term changes in the CNS induced by HIV infection rather than by active replication. Among these effects are significant changes in catecholaminergic neurotransmission, especially in dopaminergic brain regions. In HIV-infected individuals not treated with ARV show prominent neuropathology is common in dopamine-rich brain regions and altered autonomic nervous system activity. Even infected individuals on therapy, there is significant dopaminergic neuropathology, and elevated stress and norepinephrine levels correlate with a decreased effectiveness of antiretroviral drugs. As catecholamines function as immunomodulatory factors, the resultant dysregulation of catecholaminergic tone could substantially alter the development of HIVassociated neuroinflammation and neuropathology. In this review, we discuss the role of catecholamines in the etiology of HIV neuropathogenesis. Providing a comprehensive examination of what is known about these molecules in the context of HIV-associated disease demonstrates the importance of further studies in this area, and may open the door to new therapeutic strategies that specifically ameliorate the effects of catecholaminergic dysregulation on NeuroHIV.
Keywords: Catecholamines; Drug abuse; HIV; HIV-associated neurocognitive disorders; Neuroinflammation.
Copyright © 2018 Elsevier B.V. All rights reserved.
Figures
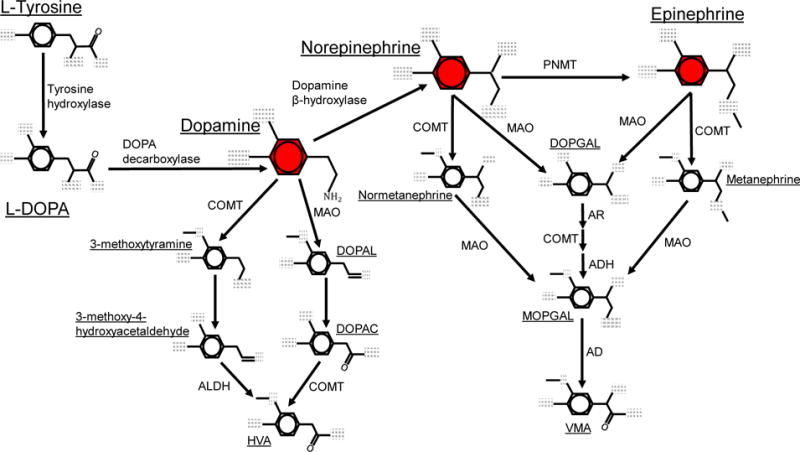
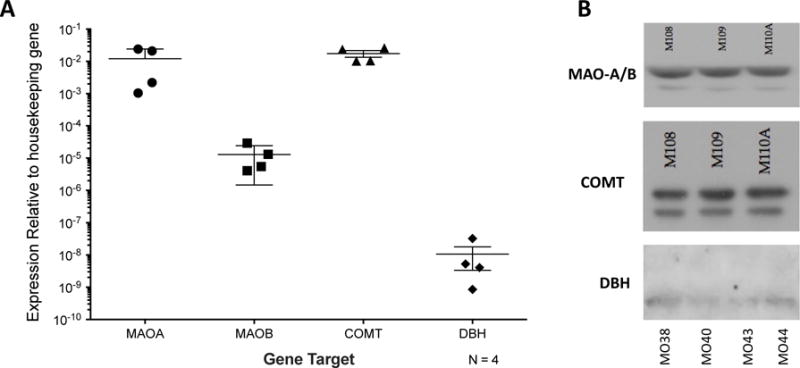
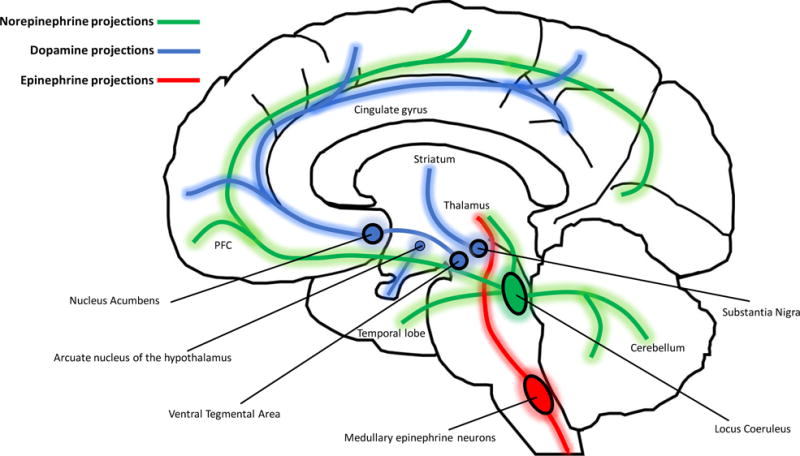
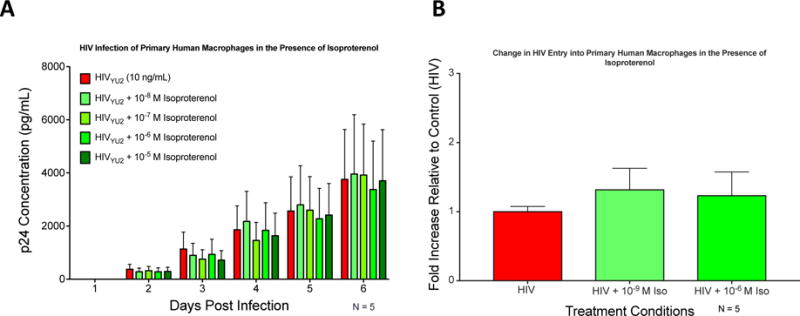
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
