

The higBA Toxin-Antitoxin Module From the Opportunistic Pathogen Acinetobacter baumannii - Regulation, Activity, and Evolution
- PMID: 29706946
- PMCID: PMC5906591
- DOI: 10.3389/fmicb.2018.00732
The higBA Toxin-Antitoxin Module From the Opportunistic Pathogen Acinetobacter baumannii - Regulation, Activity, and Evolution
Abstract
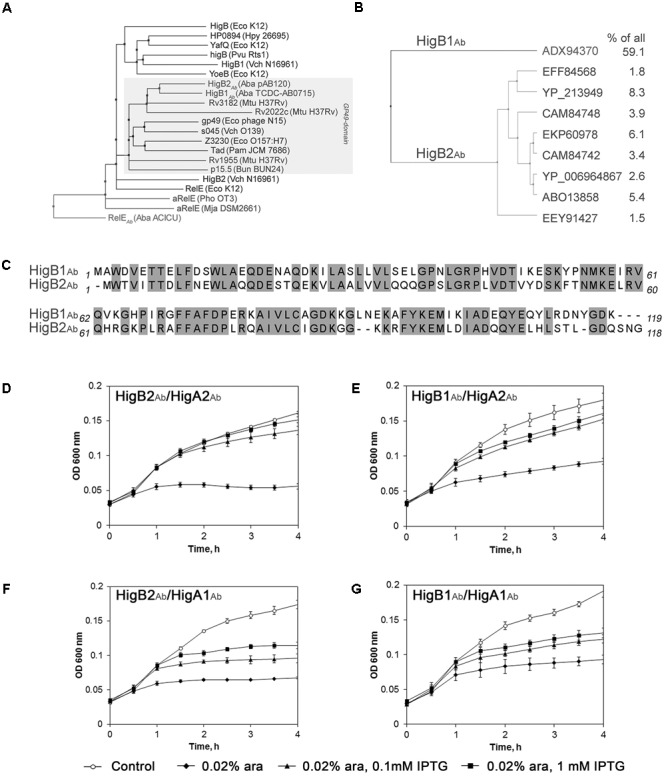
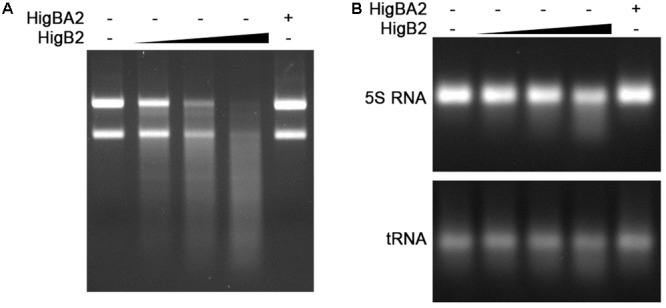
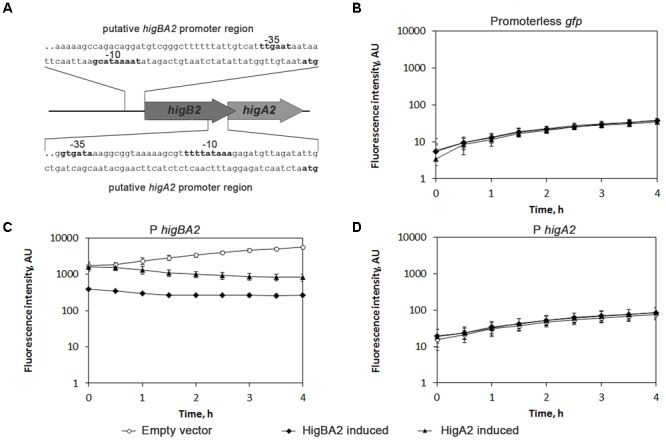
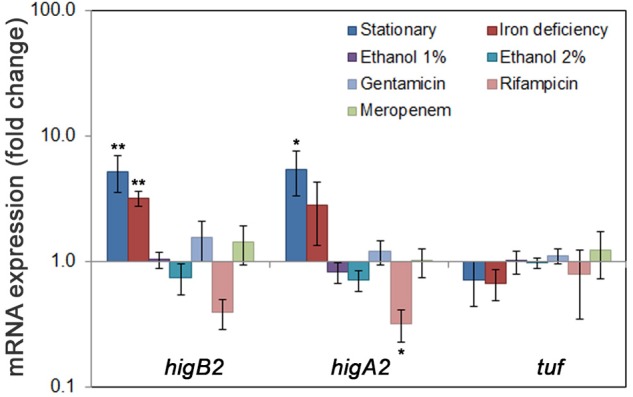
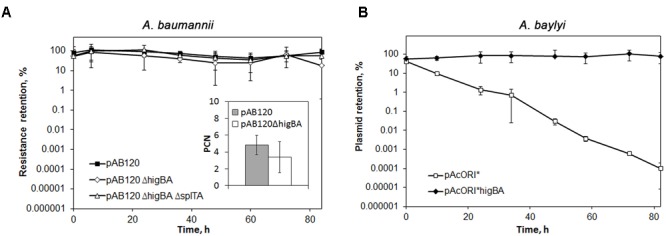
Acinetobacter baumannii is one of the major causes of hard to treat multidrug-resistant hospital infections. A. baumannii features contributing to its spread and persistence in clinical environment are only beginning to be explored. Bacterial toxin-antitoxin (TA) systems are genetic loci shown to be involved in plasmid maintenance and proposed to function as components of stress response networks. Here we present a thorough characterization of type II system of A. baumannii, which is the most ubiquitous TA module present in A. baumannii plasmids. higBA of A. baumannii is a reverse TA (the toxin gene is the first in the operon) and shows little homology to other TA systems of RelE superfamily. It is represented by two variants, which both are functional albeit exhibit strong difference in sequence conservation. The higBA2 operon is found on ubiquitous 11 Kb pAB120 plasmid, conferring carbapenem resistance to clinical A. baumannii isolates and represents a higBA variant that can be found with multiple sequence variations. We show here that higBA2 is capable to confer maintenance of unstable plasmid in Acinetobacter species. HigB2 toxin functions as a ribonuclease and its activity is neutralized by HigA2 antitoxin through formation of an unusually large heterooligomeric complex. Based on the in vivo expression analysis of gfp reporter gene we propose that HigA2 antitoxin and HigBA2 protein complex bind the higBA2 promoter region to downregulate its transcription. We also demonstrate that higBA2 is a stress responsive locus, whose transcription changes in conditions encountered by A. baumannii in clinical environment and within the host. We show elevated expression of higBA2 during stationary phase, under iron deficiency and downregulated expression after antibiotic (rifampicin) treatment.
Keywords: Acinetobacter baumannii; HigBA; plasmid maintenance; protein complex; toxin-antitoxin.
Figures

References
-
- Cárdenas-Mondragón M. G., Ares M. A., Panunzi L. G., Pacheco S., Camorlinga-Ponce M., Girón J. A., et al. (2016). Transcriptional profiling of type II toxin-antitoxin genes of Helicobacter pylori under different environmental conditions: identification of HP0967-HP0968 system. Front. Microbiol. 7:1872. 10.3389/fmicb.2016.01872 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources