

Co-translational protein targeting facilitates centrosomal recruitment of PCNT during centrosome maturation in vertebrates
- PMID: 29708497
- PMCID: PMC5976437
- DOI: 10.7554/eLife.34959
Co-translational protein targeting facilitates centrosomal recruitment of PCNT during centrosome maturation in vertebrates
Abstract
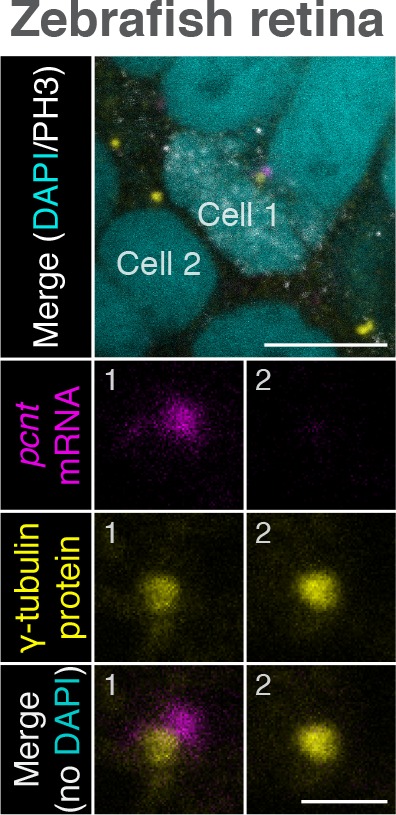
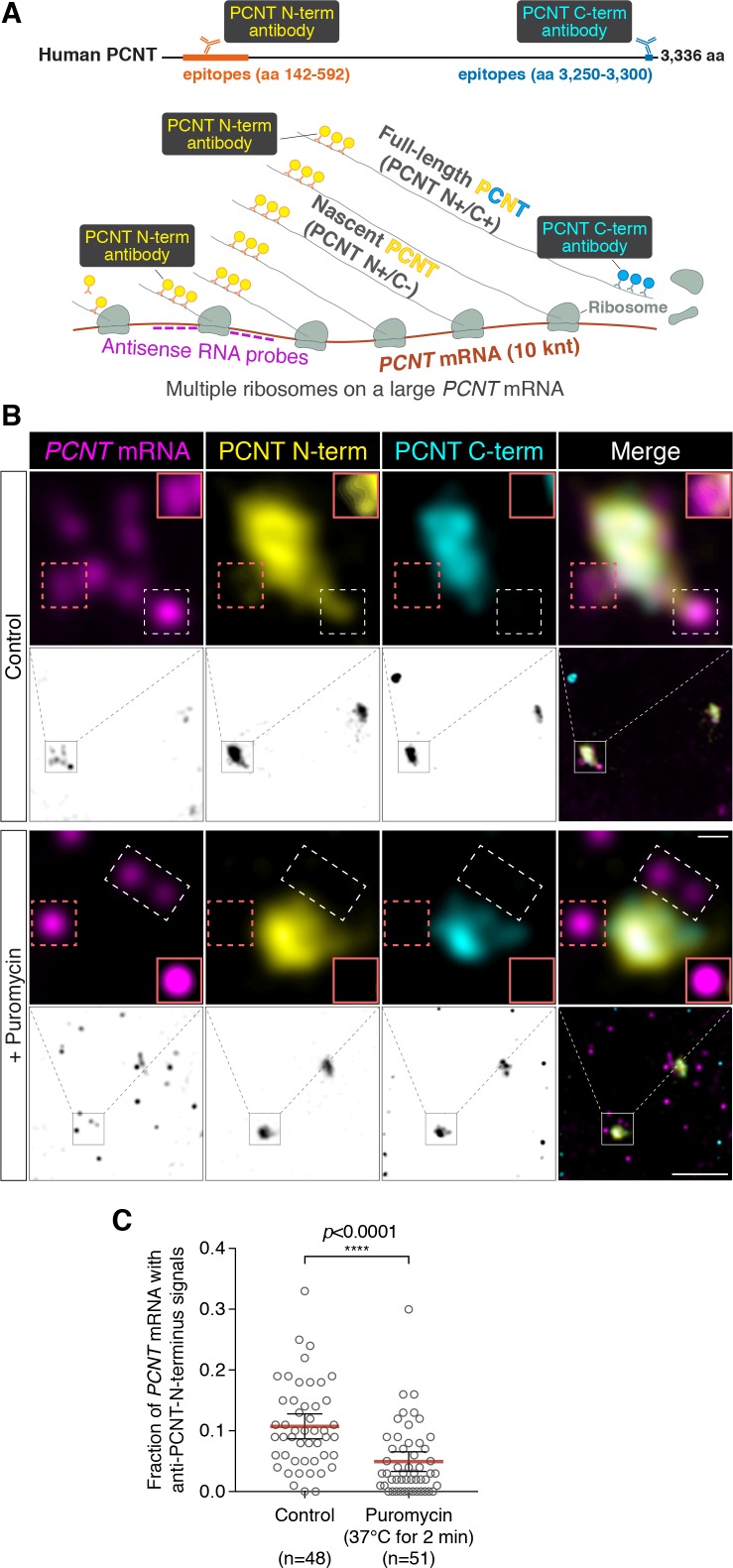
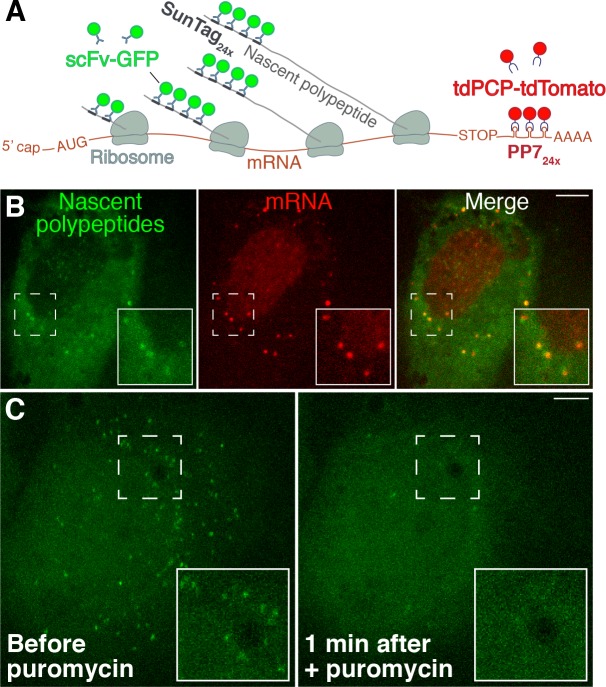
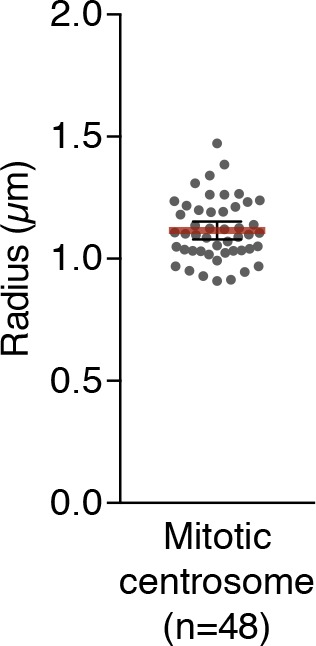
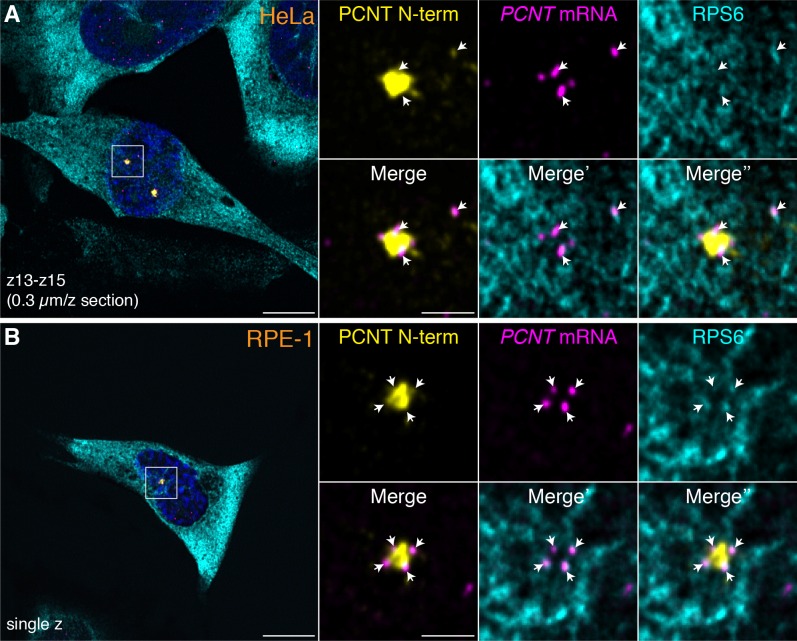
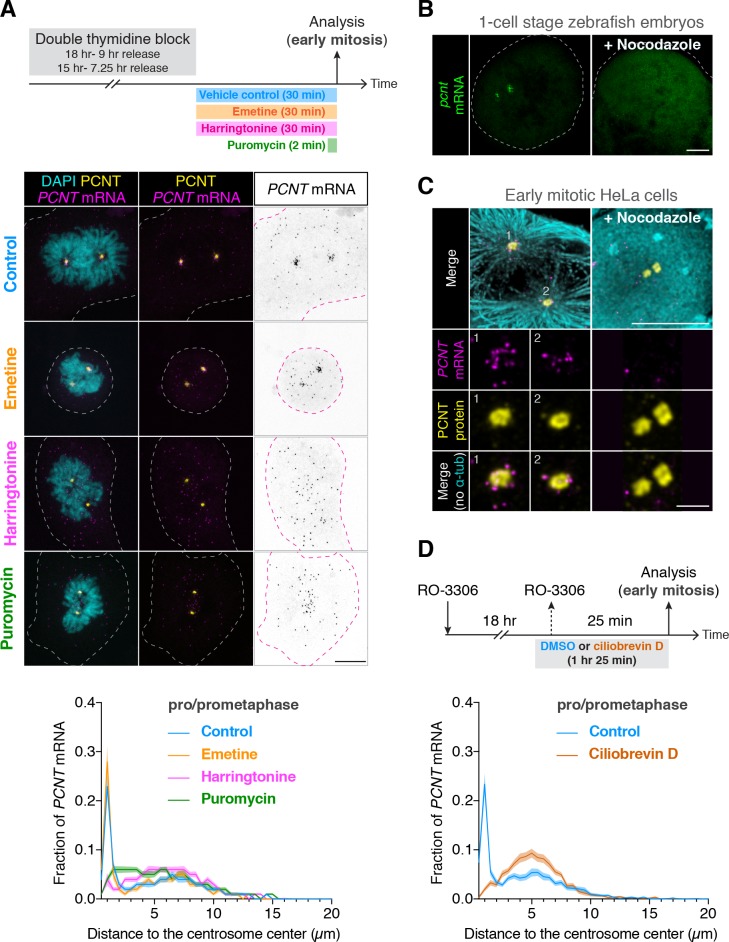
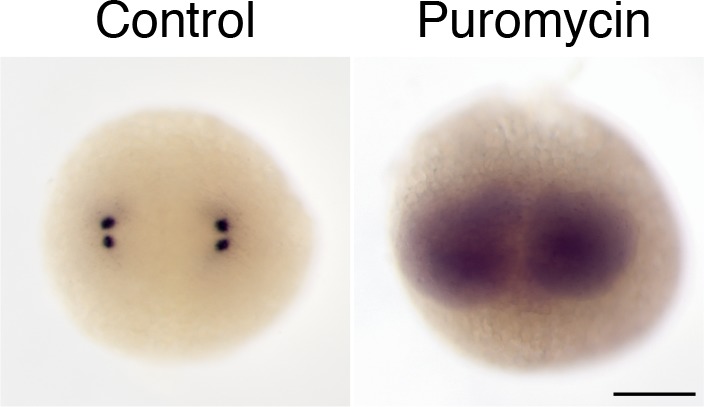
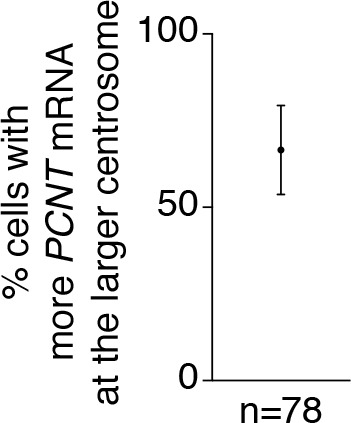
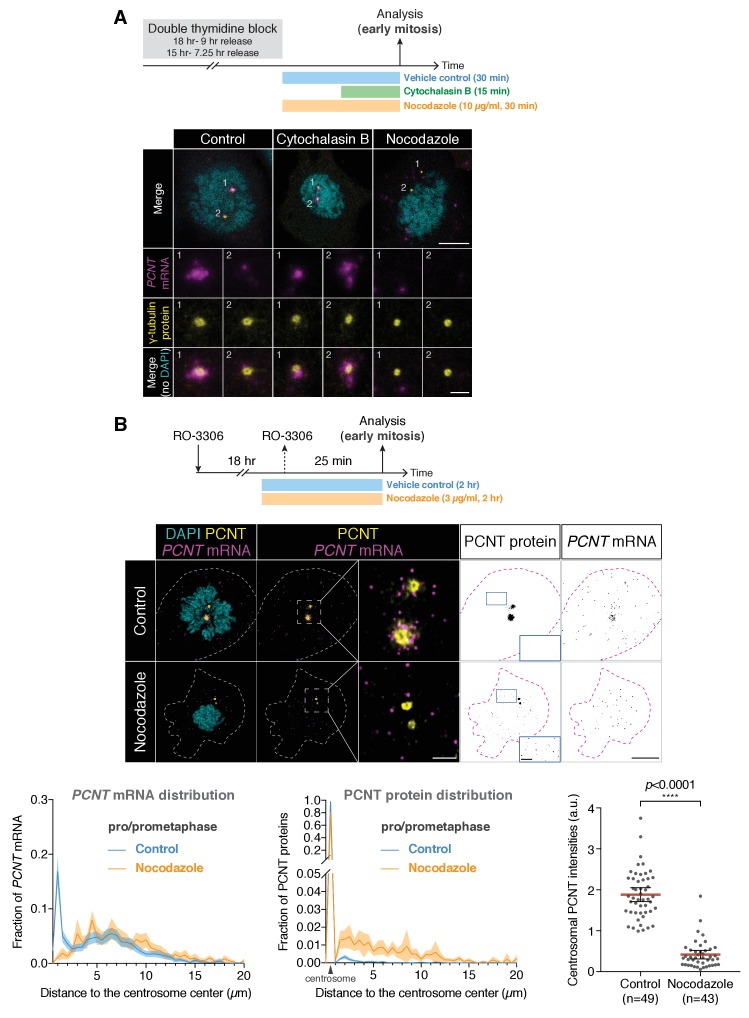
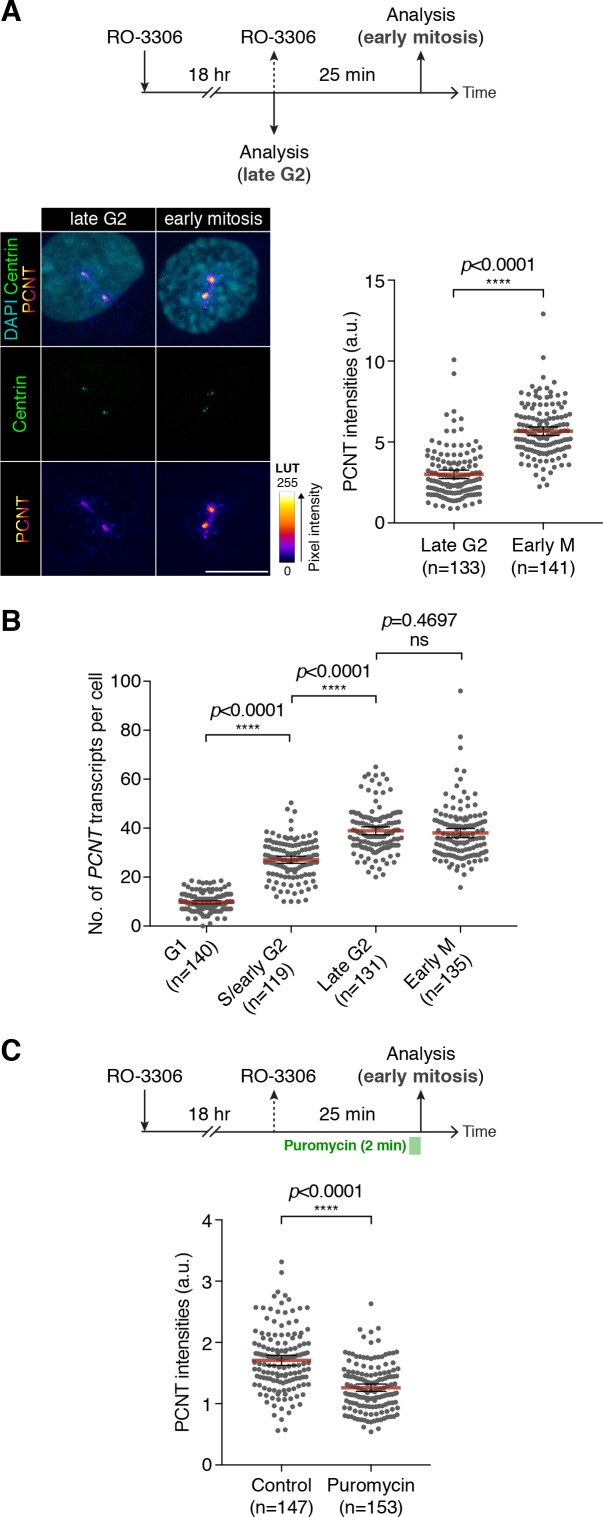
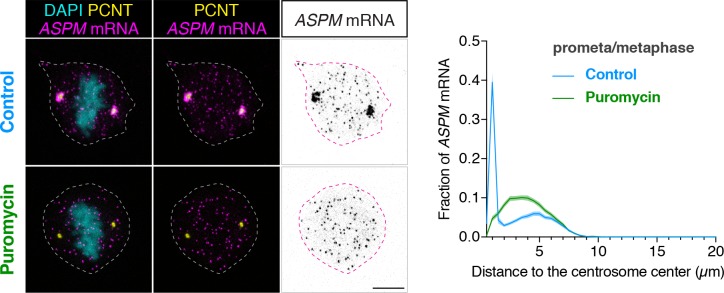
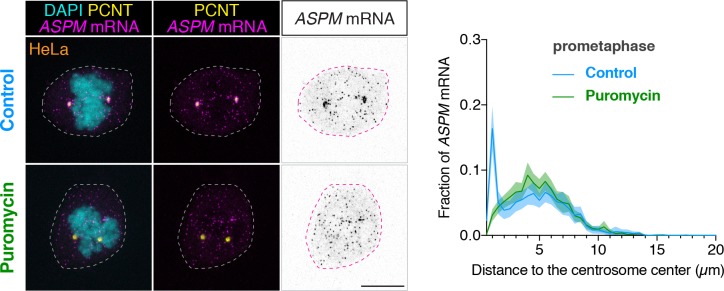
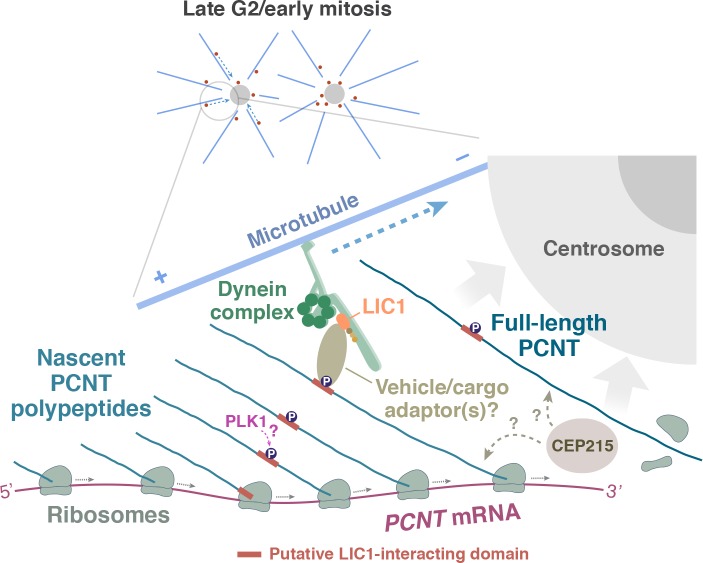
As microtubule-organizing centers of animal cells, centrosomes guide the formation of the bipolar spindle that segregates chromosomes during mitosis. At mitosis onset, centrosomes maximize microtubule-organizing activity by rapidly expanding the pericentriolar material (PCM). This process is in part driven by the large PCM protein pericentrin (PCNT), as its level increases at the PCM and helps recruit additional PCM components. However, the mechanism underlying the timely centrosomal enrichment of PCNT remains unclear. Here, we show that PCNT is delivered co-translationally to centrosomes during early mitosis by cytoplasmic dynein, as evidenced by centrosomal enrichment of PCNT mRNA, its translation near centrosomes, and requirement of intact polysomes for PCNT mRNA localization. Additionally, the microtubule minus-end regulator, ASPM, is also targeted co-translationally to mitotic spindle poles. Together, these findings suggest that co-translational targeting of cytoplasmic proteins to specific subcellular destinations may be a generalized protein targeting mechanism.
Keywords: cell biology; centrosome; human; mRNA localization; translation; zebrafish.
© 2018, Sepulveda et al.
Conflict of interest statement
GS, MA, IB, KM, TO, NC, LC, LC, XJ, DY, BH, LJ No competing interests declared
Figures
Similar articles
-
PLK1 phosphorylation of pericentrin initiates centrosome maturation at the onset of mitosis.J Cell Biol. 2011 Dec 26;195(7):1093-101. doi: 10.1083/jcb.201106093. Epub 2011 Dec 19. J Cell Biol. 2011. PMID: 22184200 Free PMC article.
-
PCNT is critical for the association and conversion of centrioles to centrosomes during mitosis.J Cell Sci. 2019 Mar 26;132(6):jcs225789. doi: 10.1242/jcs.225789. J Cell Sci. 2019. PMID: 30814333
-
CEP215 is involved in the dynein-dependent accumulation of pericentriolar matrix proteins for spindle pole formation.Cell Cycle. 2010 Feb 15;9(4):774-83. Epub 2010 Feb 9. Cell Cycle. 2010. PMID: 20139723
-
Construction of centrosomes and spindle poles by molecular motor-driven assembly of protein particles.Traffic. 2000 Dec;1(12):927-34. Traffic. 2000. PMID: 11208082 Review.
-
Polar expeditions--provisioning the centrosome for mitosis.Nat Cell Biol. 2003 Jun;5(6):505-11. doi: 10.1038/ncb0603-505. Nat Cell Biol. 2003. PMID: 12776127 Review.
Cited by
-
Radial glia progenitor polarity in health and disease.Front Cell Dev Biol. 2024 Oct 2;12:1478283. doi: 10.3389/fcell.2024.1478283. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 39416687 Free PMC article. Review.
-
The Physiological Roles of the Exon Junction Complex in Development and Diseases.Cells. 2022 Apr 1;11(7):1192. doi: 10.3390/cells11071192. Cells. 2022. PMID: 35406756 Free PMC article. Review.
-
Specific mitotic events drive left-right organizer development.Development. 2025 May 15;152(10):dev204687. doi: 10.1242/dev.204687. Epub 2025 May 19. Development. 2025. PMID: 40230260 Free PMC article.
-
Exon junction complex dependent mRNA localization is linked to centrosome organization during ciliogenesis.Nat Commun. 2021 Mar 1;12(1):1351. doi: 10.1038/s41467-021-21590-w. Nat Commun. 2021. PMID: 33649372 Free PMC article.
-
The ERM-1 membrane-binding domain directs erm-1 mRNA localization to the plasma membrane in the C. elegans embryo.Development. 2022 Nov 15;149(22):dev200930. doi: 10.1242/dev.200930. Epub 2022 Nov 18. Development. 2022. PMID: 36314842 Free PMC article.
References
-
- Anitha A, Nakamura K, Yamada K, Iwayama Y, Toyota T, Takei N, Iwata Y, Suzuki K, Sekine Y, Matsuzaki H, Kawai M, Thanseem I, Miyoshi K, Katayama T, Matsuzaki S, Baba K, Honda A, Hattori T, Shimizu S, Kumamoto N, Kikuchi M, Tohyama M, Yoshikawa T, Mori N. Association studies and gene expression analyses of the DISC1-interacting molecules, pericentrin 2 (PCNT2) and DISC1-binding zinc finger protein (DBZ), with schizophrenia and with bipolar disorder. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics. 2009;150B:967–976. doi: 10.1002/ajmg.b.30926. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
