

Compartment-Specific Labeling of Bacterial Periplasmic Proteins by Peroxidase-Mediated Biotinylation
- PMID: 29708735
- PMCID: PMC6767932
- DOI: 10.1021/acsinfecdis.8b00044
Compartment-Specific Labeling of Bacterial Periplasmic Proteins by Peroxidase-Mediated Biotinylation
Erratum in
-
Correction to Compartment-Specific Labeling of Bacterial Periplasmic Proteins by Peroxidase-Mediated Biotinylation.ACS Infect Dis. 2018 Jul 13;4(7):1157. doi: 10.1021/acsinfecdis.8b00143. Epub 2018 Jun 25. ACS Infect Dis. 2018. PMID: 29939011 No abstract available.
Abstract
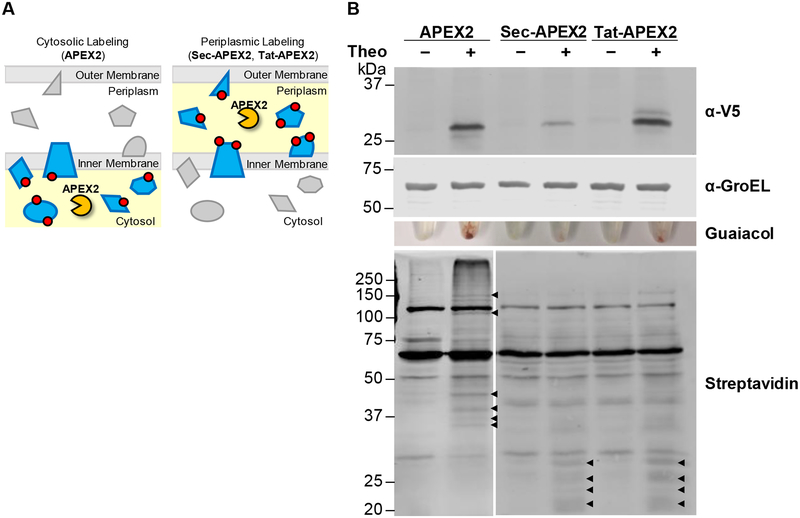
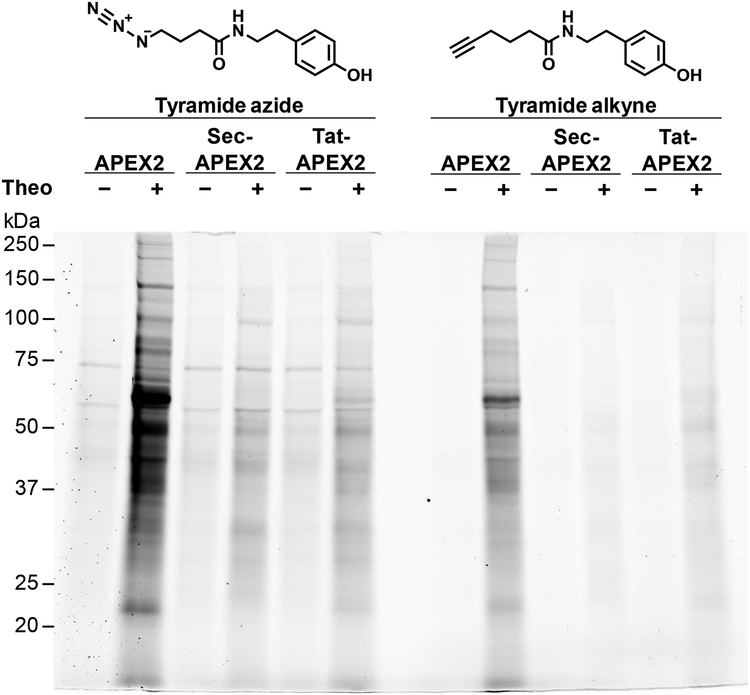
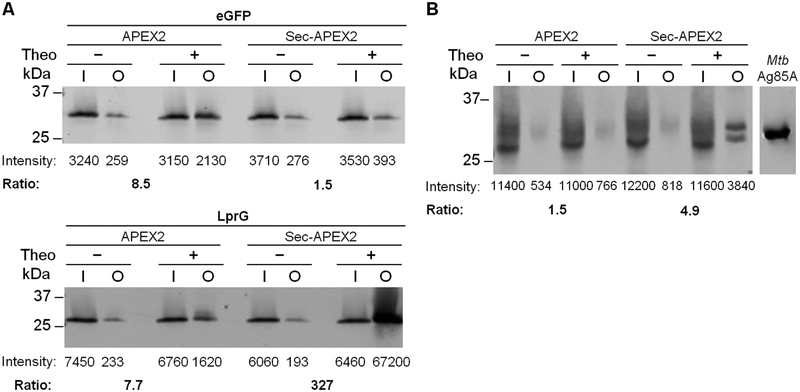
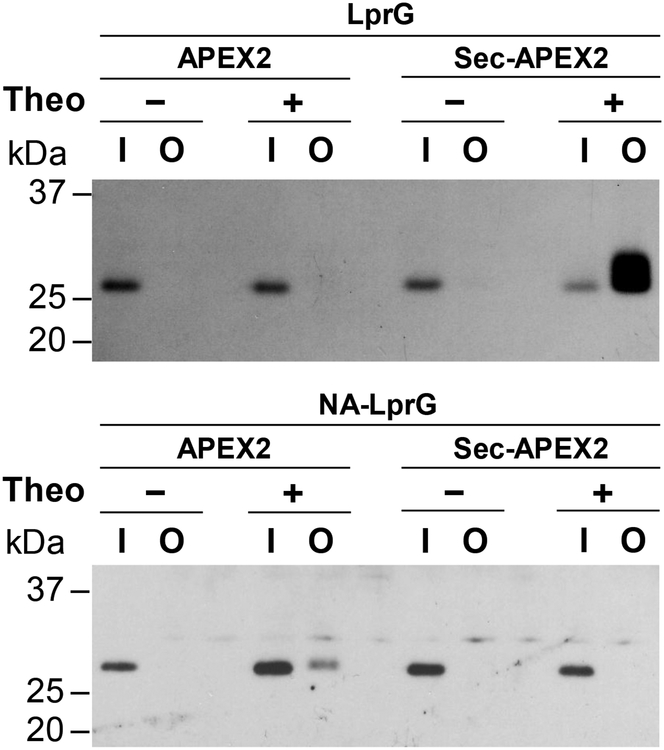
The study of the bacterial periplasm requires techniques with sufficient spatial resolution and sensitivity to resolve the components and processes within this subcellular compartment. Peroxidase-mediated biotinylation has enabled targeted labeling of proteins within subcellular compartments of mammalian cells. We investigated whether this methodology could be applied to the bacterial periplasm. In this study, we demonstrated that peroxidase-mediated biotinylation can be performed in mycobacteria and Escherichia coli. To eliminate detection artifacts from natively biotinylated mycobacterial proteins, we validated two alternative labeling substrates, tyramide azide and tyramide alkyne, which enable biotin-independent detection of labeled proteins. We also targeted peroxidase expression to the periplasm, resulting in compartment-specific labeling of periplasmic versus cytoplasmic proteins in mycobacteria. Finally, we showed that this method can be used to validate protein relocalization to the cytoplasm upon removal of a secretion signal. This novel application of peroxidase-mediated protein labeling will advance efforts to characterize the role of the periplasm in bacterial physiology and pathogenesis.
Keywords: APEX; CuAAC click chemistry; bacteria; mycobacteria; periplasm; peroxidase-mediated biotinylation.
Figures
References
-
- Seeliger JC; Holsclaw CM; Schelle MW; Botyanszki Z; Gilmore SA; Tully SE; Niederweis M; Cravatt BF; Leary JA; Bertozzi CR, Elucidation and chemical modulation of sulfolipid-1 biosynthesis in Mycobacterium tuberculosis. J. Biol. Chem 2012, 287 (11), 7990–8000. DOI: 10.1074/jbc.M111.315473. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
