

Escherichia coli cultures maintain stable subpopulation structure during long-term evolution
- PMID: 29712844
- PMCID: PMC5960275
- DOI: 10.1073/pnas.1708371115
Escherichia coli cultures maintain stable subpopulation structure during long-term evolution
Abstract
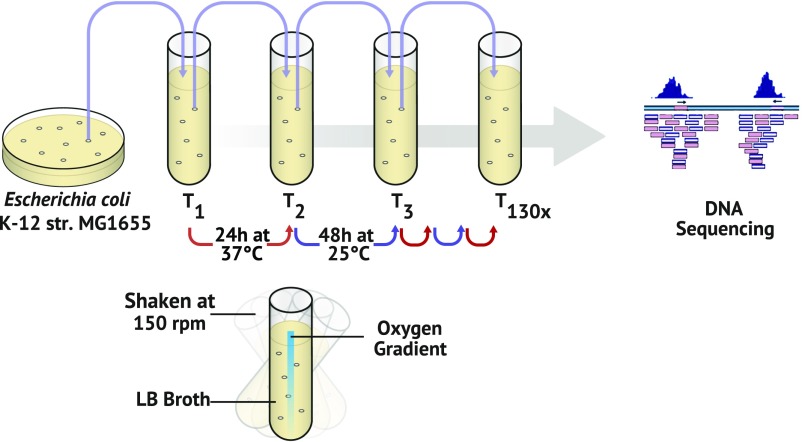
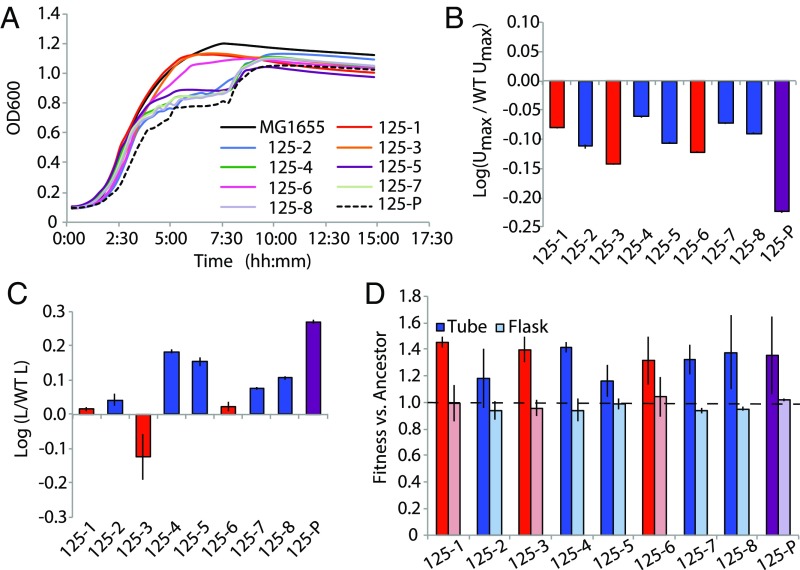
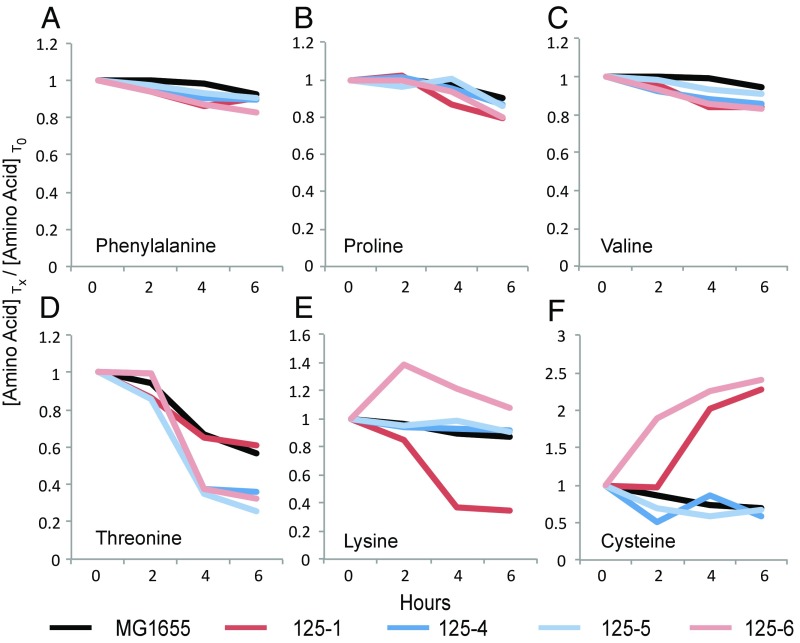
How genetic variation is generated and maintained remains a central question in evolutionary biology. When presented with a complex environment, microbes can take advantage of genetic variation to exploit new niches. Here we present a massively parallel experiment where WT and repair-deficient (∆mutL) Escherichia coli populations have evolved over 3 y in a spatially heterogeneous and nutritionally complex environment. Metagenomic sequencing revealed that these initially isogenic populations evolved and maintained stable subpopulation structure in just 10 mL of medium for up to 10,000 generations, consisting of up to five major haplotypes with many minor haplotypes. We characterized the genomic, transcriptomic, exometabolomic, and phenotypic differences between clonal isolates, revealing subpopulation structure driven primarily by spatial segregation followed by differential utilization of nutrients. In addition to genes regulating the import and catabolism of nutrients, major polymorphisms of note included insertion elements transposing into fimE (regulator of the type I fimbriae) and upstream of hns (global regulator of environmental-change and stress-response genes), both known to regulate biofilm formation. Interestingly, these genes have also been identified as critical to colonization in uropathogenic E. coli infections. Our findings illustrate the complexity that can arise and persist even in small cultures, raising the possibility that infections may often be promoted by an evolving and complex pathogen population.
Keywords: biofilm; complex environment; niche specialization.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Friesen ML, Saxer G, Travisano M, Doebeli M. Experimental evidence for sympatric ecological diversification due to frequency-dependent competition in Escherichia coli. Evolution. 2004;58:245–260. - PubMed
-
- Rozen DE, Philippe N, Arjan de Visser J, Lenski RE, Schneider D. Death and cannibalism in a seasonal environment facilitate bacterial coexistence. Ecol Lett. 2009;12:34–44. - PubMed
-
- Rainey PB, Travisano M. Adaptive radiation in a heterogeneous environment. Nature. 1998;394:69–72. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
