

Callosal Influence on Visual Receptive Fields Has an Ocular, an Orientation-and Direction Bias
- PMID: 29713267
- PMCID: PMC5911488
- DOI: 10.3389/fnsys.2018.00011
Callosal Influence on Visual Receptive Fields Has an Ocular, an Orientation-and Direction Bias
Abstract
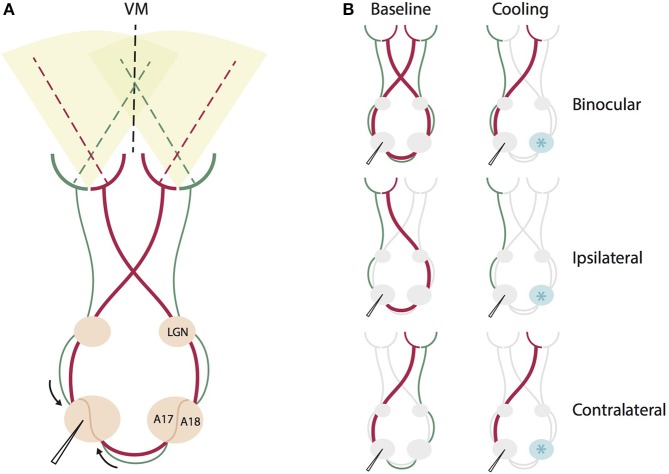
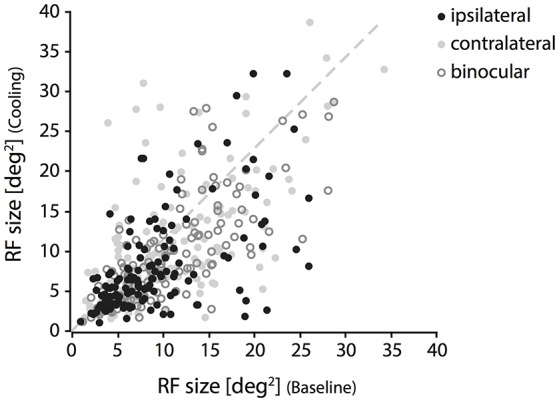
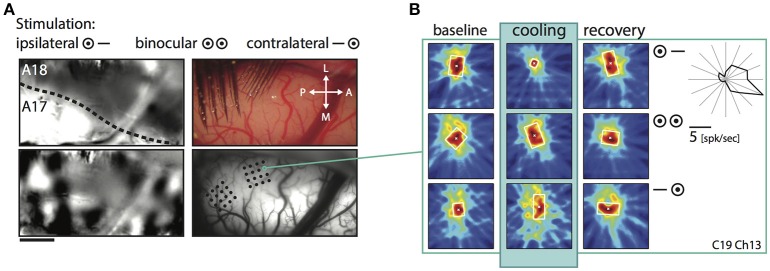
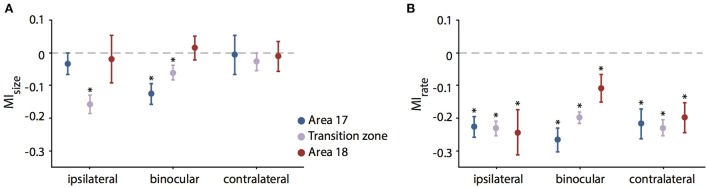
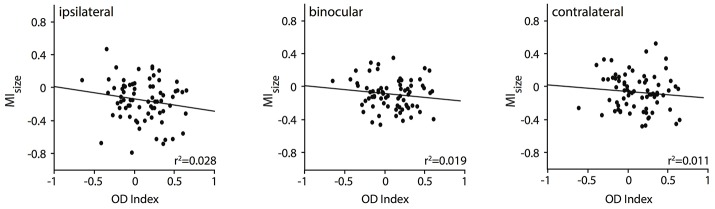
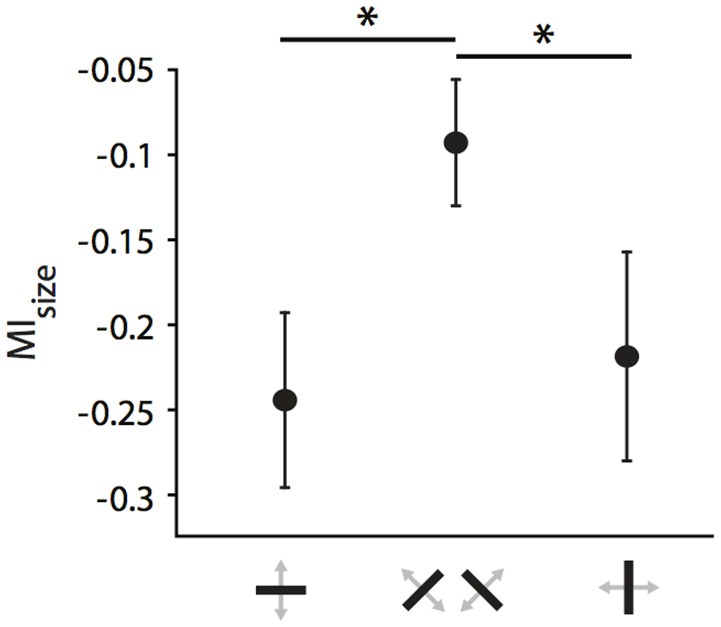
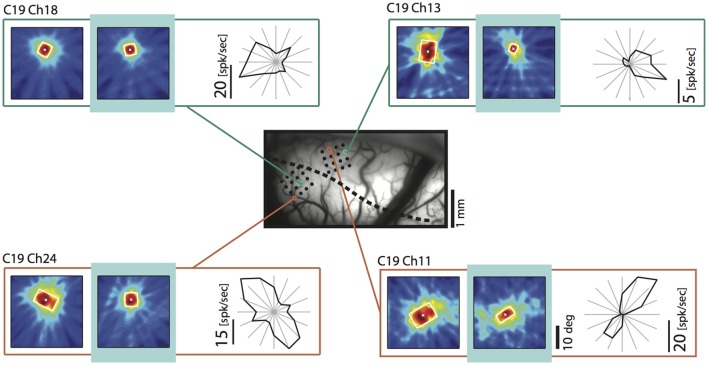
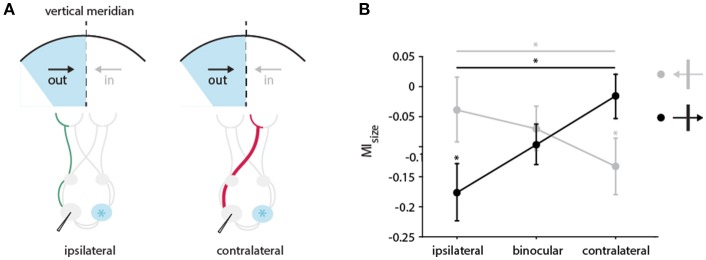
One leading hypothesis on the nature of visual callosal connections (CC) is that they replicate features of intrahemispheric lateral connections. However, CC act also in the central part of the binocular visual field. In agreement, early experiments in cats indicated that they provide the ipsilateral eye part of binocular receptive fields (RFs) at the vertical midline (Berlucchi and Rizzolatti, 1968), and play a key role in stereoscopic function. But until today callosal inputs to receptive fields activated by one or both eyes were never compared simultaneously, because callosal function has been often studied by cutting or lesioning either corpus callosum or optic chiasm not allowing such a comparison. To investigate the functional contribution of CC in the intact cat visual system we recorded both monocular and binocular neuronal spiking responses and receptive fields in the 17/18 transition zone during reversible deactivation of the contralateral hemisphere. Unexpectedly from many of the previous reports, we observe no change in ocular dominance during CC deactivation. Throughout the transition zone, a majority of RFs shrink, but several also increase in size. RFs are significantly more affected for ipsi- as opposed to contralateral stimulation, but changes are also observed with binocular stimulation. Noteworthy, RF shrinkages are tiny and not correlated to the profound decreases of monocular and binocular firing rates. They depend more on orientation and direction preference than on eccentricity or ocular dominance of the receiving neuron's RF. Our findings confirm that in binocularly viewing mammals, binocular RFs near the midline are constructed via the direct geniculo-cortical pathway. They also support the idea that input from the two eyes complement each other through CC: Rather than linking parts of RFs separated by the vertical meridian, CC convey a modulatory influence, reflecting the feature selectivity of lateral circuits, with a strong cardinal bias.
Keywords: anticipation; binocular; interhemispheric connectivity; monocular; orientation selectivity.
Figures

Similar articles
-
Unilateral paralytic strabismus in the adult cat induces plastic changes in interocular disparity along the visual midline: contribution of the corpus callosum.Vis Neurosci. 2005 May-Jun;22(3):325-43. doi: 10.1017/S0952523805223088. Vis Neurosci. 2005. PMID: 16079008
-
Importance of corpus callosum for visual receptive fields of single neurons in cat superior colliculus.J Neurophysiol. 1979 Jan;42(1 Pt 1):137-52. doi: 10.1152/jn.1979.42.1.137. J Neurophysiol. 1979. PMID: 430108
-
Aberrant visual projections in the Siamese cat.J Physiol. 1971 Oct;218(1):33-62. doi: 10.1113/jphysiol.1971.sp009603. J Physiol. 1971. PMID: 5130620 Free PMC article.
-
The deafferented visual cortex and interhemispheric relationships: a physiological approach.Metab Pediatr Syst Ophthalmol (1985). 1988;11(1-2):100-10. Metab Pediatr Syst Ophthalmol (1985). 1988. PMID: 3076606 Review.
-
Cortical plasticity revealed by circumscribed retinal lesions or artificial scotomas.Prog Brain Res. 2001;134:217-46. doi: 10.1016/s0079-6123(01)34016-5. Prog Brain Res. 2001. PMID: 11702546 Review.
Cited by
-
A frontal transcallosal inhibition loop mediates interhemispheric balance in visuospatial processing.Nat Commun. 2023 Aug 25;14(1):5213. doi: 10.1038/s41467-023-40985-5. Nat Commun. 2023. PMID: 37626171 Free PMC article.
-
Recurrent dynamics in the cerebral cortex: Integration of sensory evidence with stored knowledge.Proc Natl Acad Sci U S A. 2021 Aug 17;118(33):e2101043118. doi: 10.1073/pnas.2101043118. Proc Natl Acad Sci U S A. 2021. PMID: 34362837 Free PMC article. Review.
-
Ocular dominance columns in V1 are more susceptible than associated callosal patches to imbalance of eye input during precritical and critical periods.J Comp Neurol. 2021 Aug 1;529(11):2883-2910. doi: 10.1002/cne.25134. Epub 2021 Mar 17. J Comp Neurol. 2021. PMID: 33683706 Free PMC article.
-
Spatial clustering of orientation preference in primary visual cortex of the large rodent agouti.iScience. 2020 Dec 1;24(1):101882. doi: 10.1016/j.isci.2020.101882. eCollection 2021 Jan 22. iScience. 2020. PMID: 33354663 Free PMC article.
-
The functional characterization of callosal connections.Prog Neurobiol. 2022 Jan;208:102186. doi: 10.1016/j.pneurobio.2021.102186. Epub 2021 Nov 12. Prog Neurobiol. 2022. PMID: 34780864 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous