

Proanthocyanidins in seed coat tegmen and endospermic cap inhibit seed germination in Sapium sebiferum
- PMID: 29713566
- PMCID: PMC5924686
- DOI: 10.7717/peerj.4690
Proanthocyanidins in seed coat tegmen and endospermic cap inhibit seed germination in Sapium sebiferum
Abstract
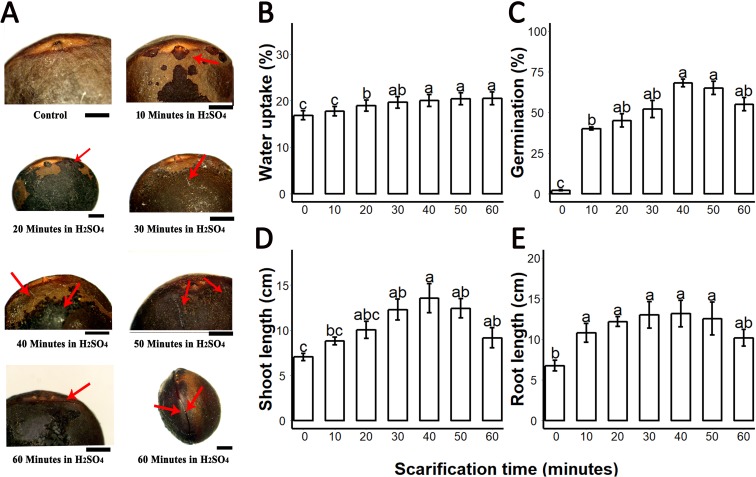
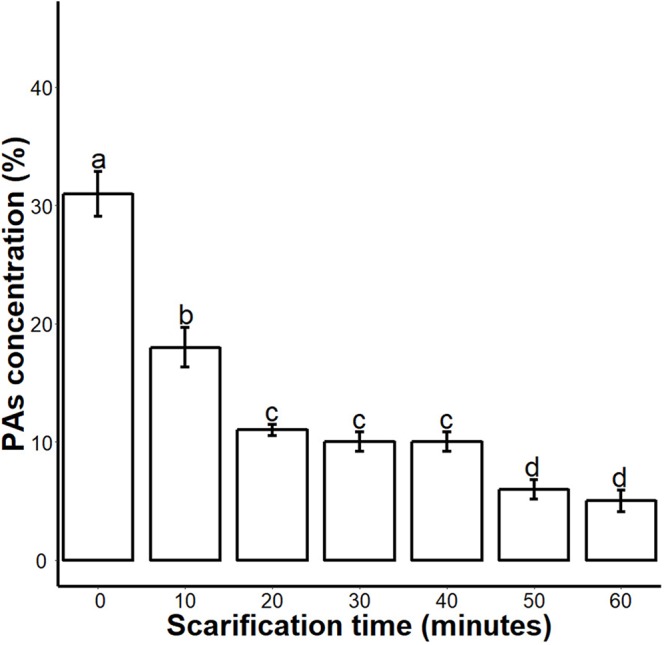
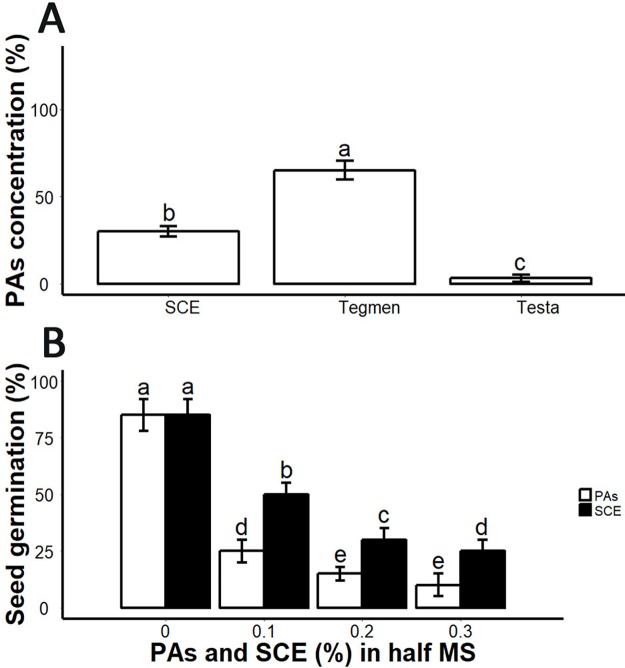
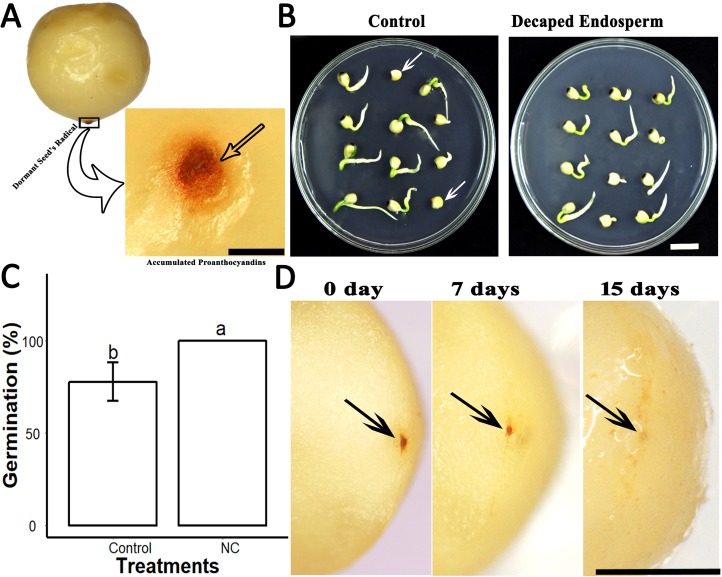
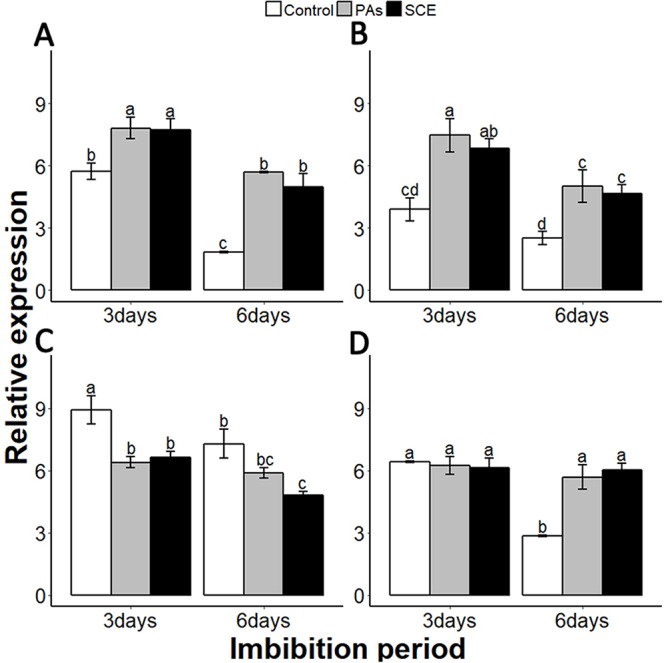
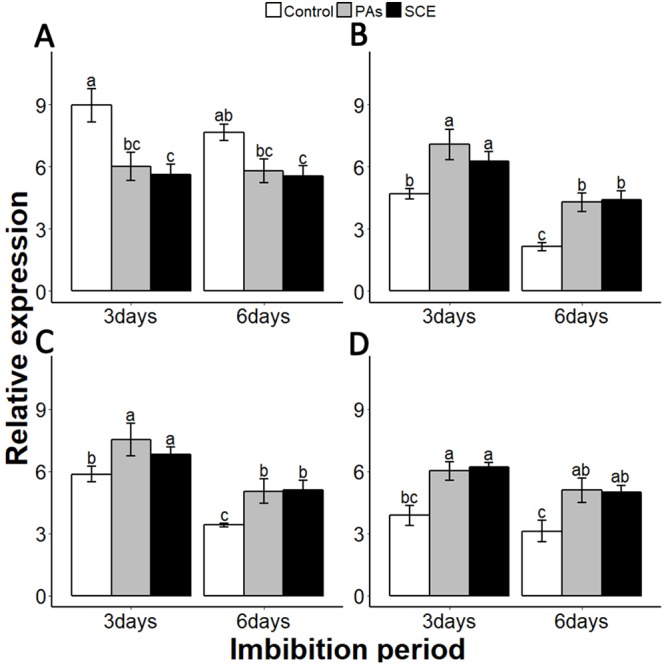
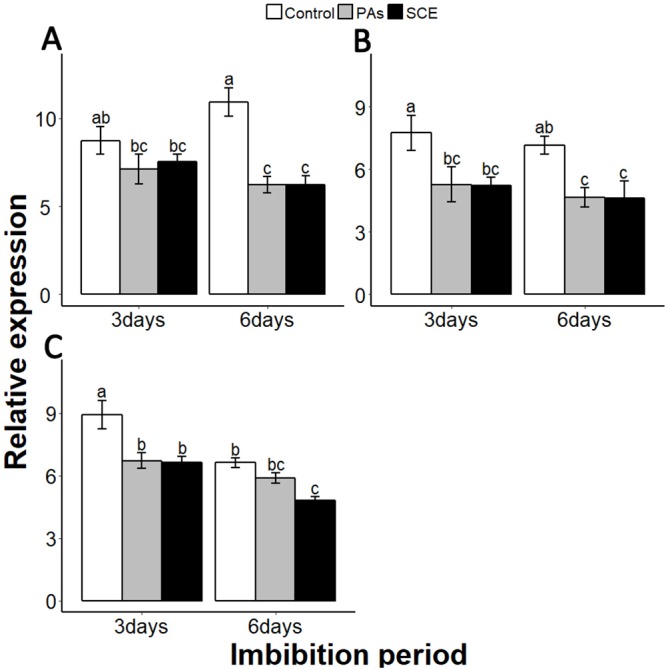
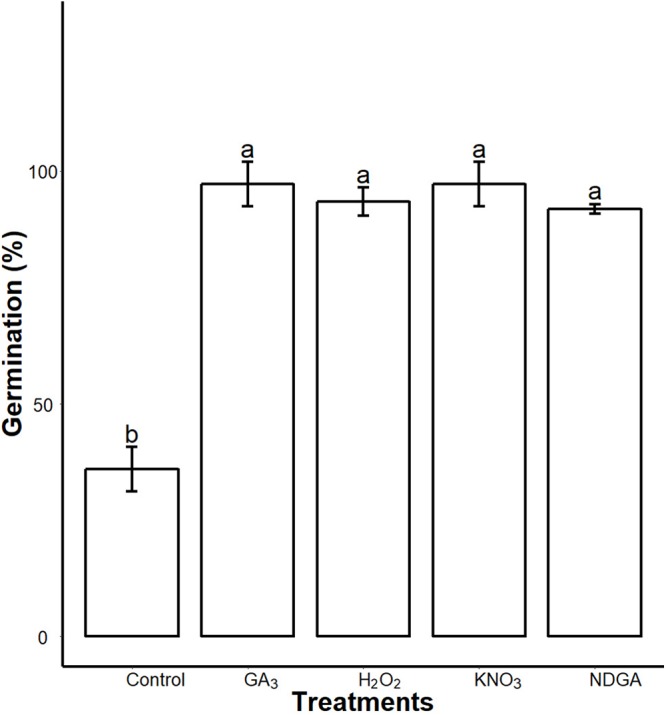
Sapium sebiferum, an ornamental and bio-energetic plant, is propagated by seed. Its seed coat contains germination inhibitors and takes a long time to stratify for germination. In this study, we discovered that the S. sebiferum seed coat (especially the tegmen) and endospermic cap (ESC) contained high levels of proanthocyanidins (PAs). Seed coat and ESC removal induced seed germination, whereas exogenous application with seed coat extract (SCE) or PAs significantly inhibited this process, suggesting that PAs in the seed coat played a major role in regulating seed germination in S. sebiferum. We further investigated how SCE affected the expression of the seed-germination-related genes. The results showed that treatment with SCE upregulated the transcription level of the dormancy-related gene, gibberellins (GAs) suppressing genes, abscisic acid (ABA) biosynthesis and signalling genes. SCE decreased the transcript levels of ABA catabolic genes, GAs biosynthesis genes, reactive oxygen species genes and nitrates-signalling genes. Exogenous application of nordihydroguaiaretic acid, gibberellic acid, hydrogen peroxide and potassium nitrate recovered seed germination in seed-coat-extract supplemented medium. In this study, we highlighted the role of PAs, and their interactions with the other germination regulators, in the regulation of seed dormancy in S. sebiferum.
Keywords: ABA; Endospermic cap; GA; Proanthocyanidins; Sapium sebiferum; Seed dormancy; Tegmen.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Baskin C, Baskin JM. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. San Diego: Academic Press; 2014.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
