

Comparative genomics of the nonlegume Parasponia reveals insights into evolution of nitrogen-fixing rhizobium symbioses
- PMID: 29717040
- PMCID: PMC5960304
- DOI: 10.1073/pnas.1721395115
Comparative genomics of the nonlegume Parasponia reveals insights into evolution of nitrogen-fixing rhizobium symbioses
Abstract
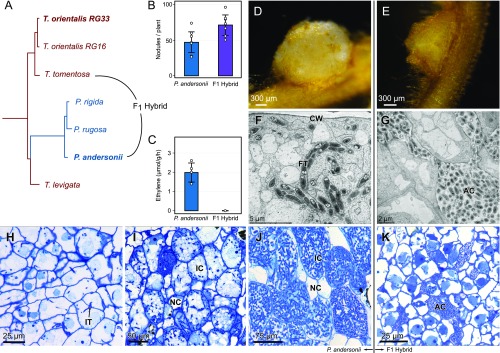
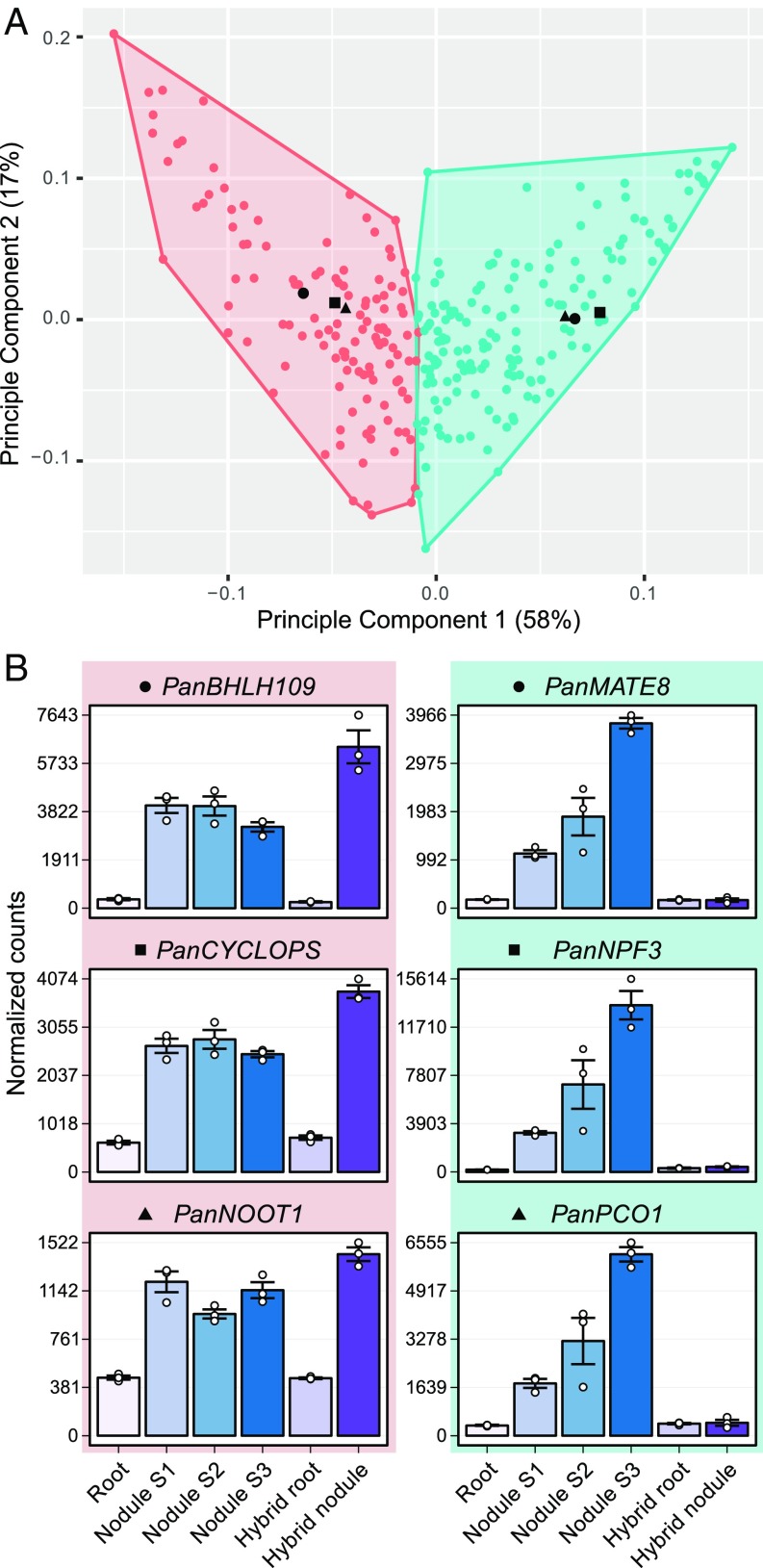
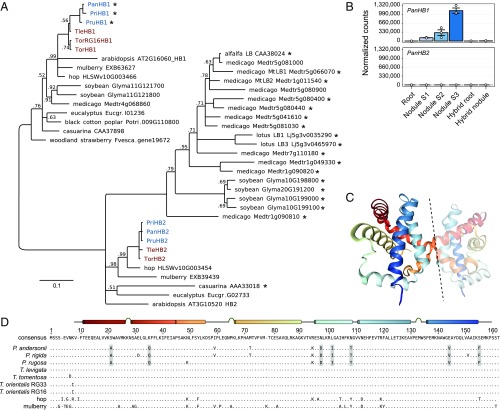
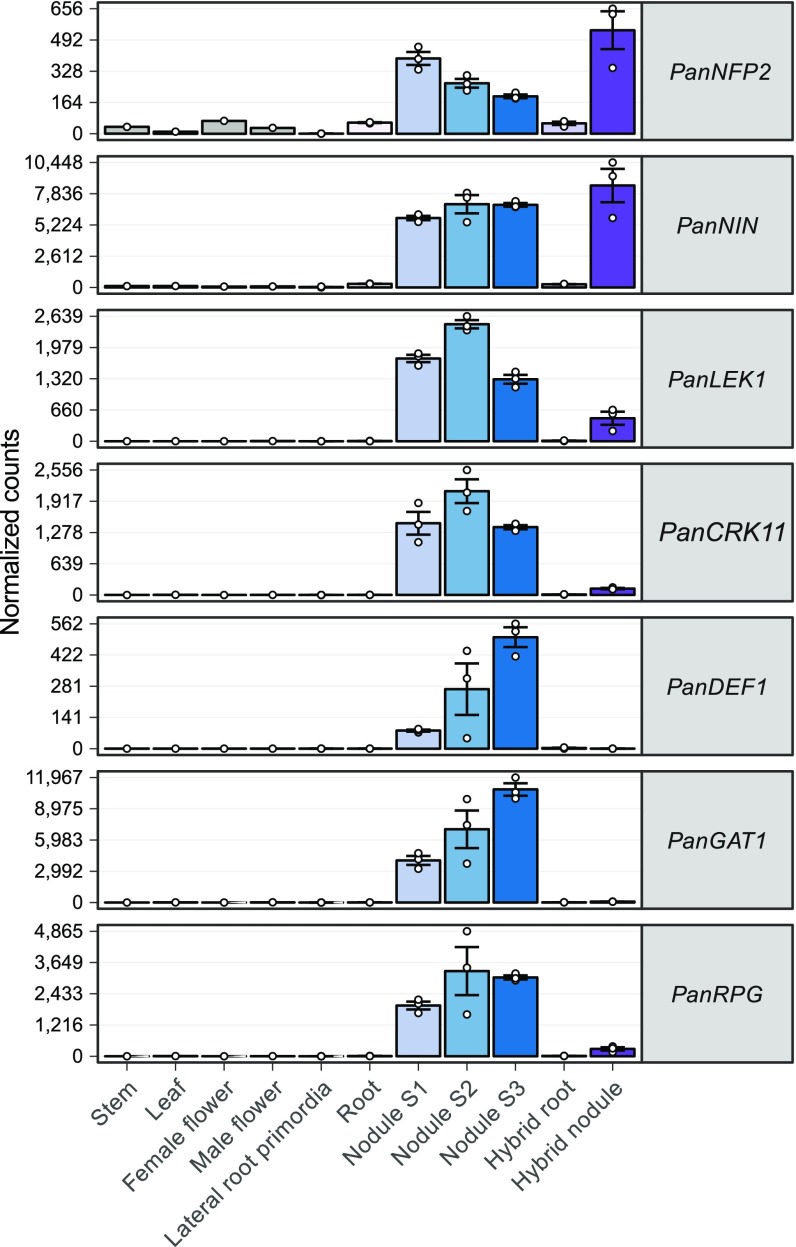
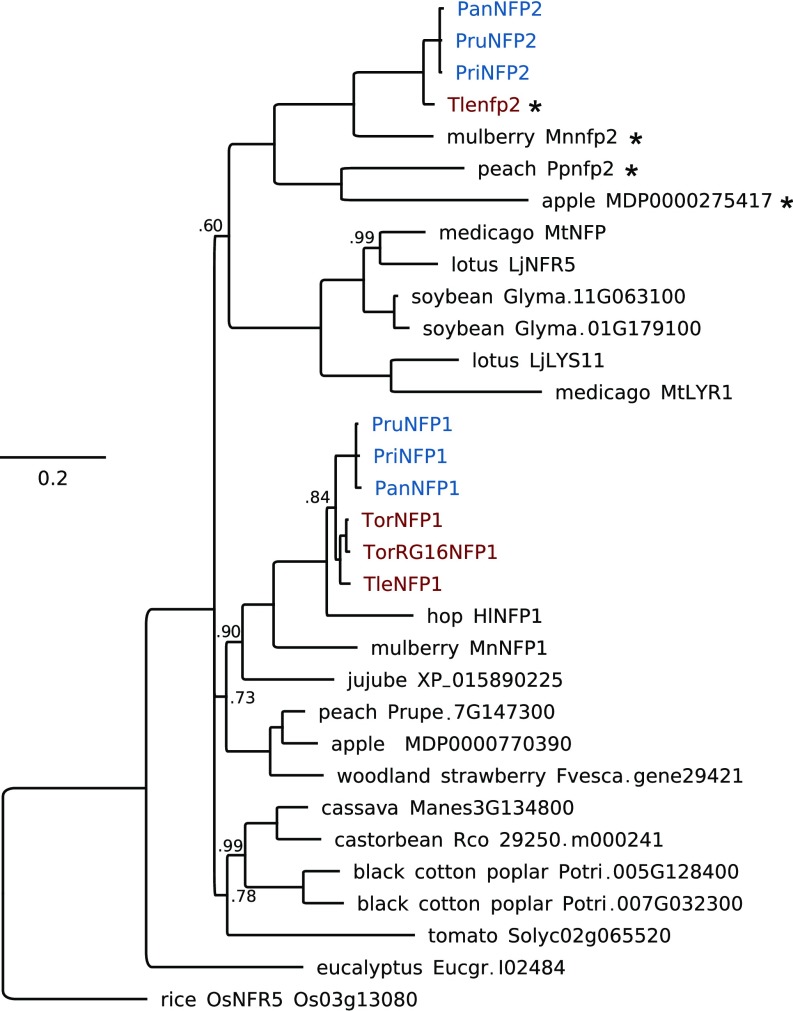
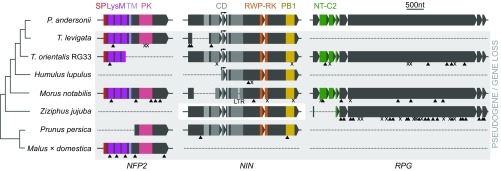
Nodules harboring nitrogen-fixing rhizobia are a well-known trait of legumes, but nodules also occur in other plant lineages, with rhizobia or the actinomycete Frankia as microsymbiont. It is generally assumed that nodulation evolved independently multiple times. However, molecular-genetic support for this hypothesis is lacking, as the genetic changes underlying nodule evolution remain elusive. We conducted genetic and comparative genomics studies by using Parasponia species (Cannabaceae), the only nonlegumes that can establish nitrogen-fixing nodules with rhizobium. Intergeneric crosses between Parasponia andersonii and its nonnodulating relative Trema tomentosa demonstrated that nodule organogenesis, but not intracellular infection, is a dominant genetic trait. Comparative transcriptomics of P. andersonii and the legume Medicago truncatula revealed utilization of at least 290 orthologous symbiosis genes in nodules. Among these are key genes that, in legumes, are essential for nodulation, including NODULE INCEPTION (NIN) and RHIZOBIUM-DIRECTED POLAR GROWTH (RPG). Comparative analysis of genomes from three Parasponia species and related nonnodulating plant species show evidence of parallel loss in nonnodulating species of putative orthologs of NIN, RPG, and NOD FACTOR PERCEPTION Parallel loss of these symbiosis genes indicates that these nonnodulating lineages lost the potential to nodulate. Taken together, our results challenge the view that nodulation evolved in parallel and raises the possibility that nodulation originated ∼100 Mya in a common ancestor of all nodulating plant species, but was subsequently lost in many descendant lineages. This will have profound implications for translational approaches aimed at engineering nitrogen-fixing nodules in crop plants.
Keywords: biological nitrogen fixation; comparative genomics; copy number variation; evolution; symbiosis.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Udvardi M, Poole PS. Transport and metabolism in legume-rhizobia symbioses. Annu Rev Plant Biol. 2013;64:781–805. - PubMed
-
- Ott T, et al. Symbiotic leghemoglobins are crucial for nitrogen fixation in legume root nodules but not for general plant growth and development. Curr Biol. 2005;15:531–535. - PubMed
-
- Burrill TJ, Hansen R. Is symbiosis possible between legume bacteria and non-legume plants? Bull Univ Ill Agric Expt Stn. 1917;202:115–181.
-
- Stokstad E. The nitrogen fix. Science. 2016;353:1225–1227. - PubMed
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
