

The stage of seed development influences iron bioavailability in pea (Pisum sativum L.)
- PMID: 29720667
- PMCID: PMC5932076
- DOI: 10.1038/s41598-018-25130-3
The stage of seed development influences iron bioavailability in pea (Pisum sativum L.)
Abstract
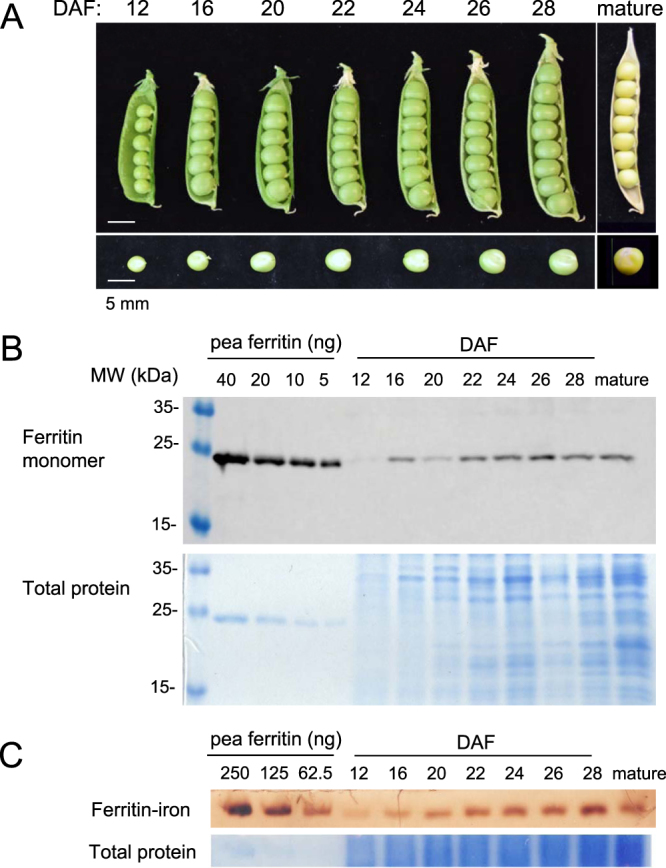
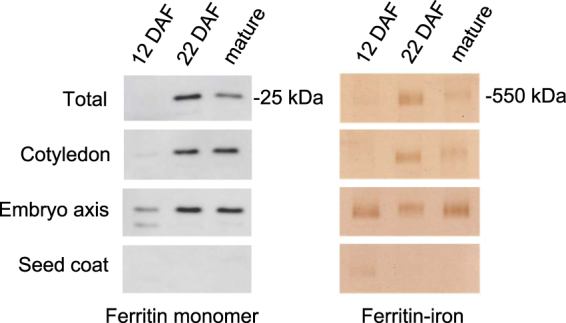
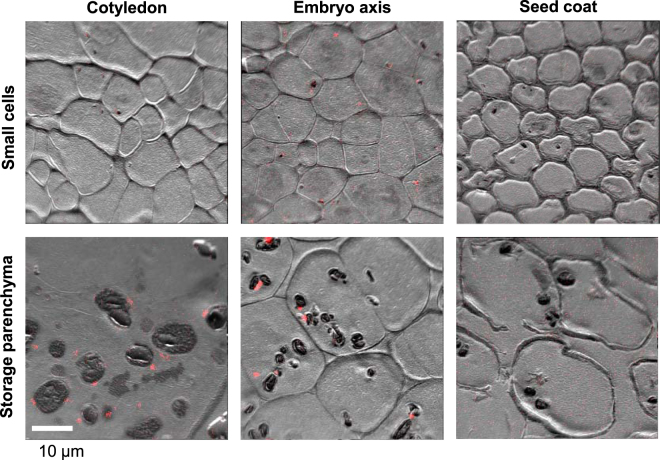
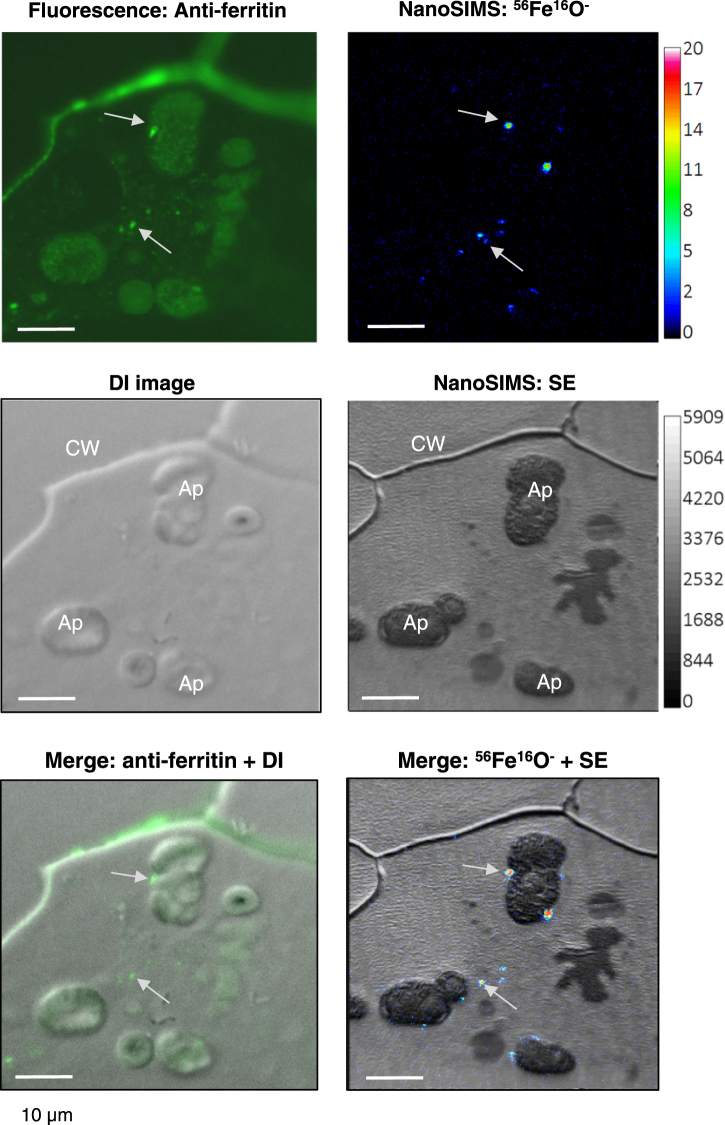
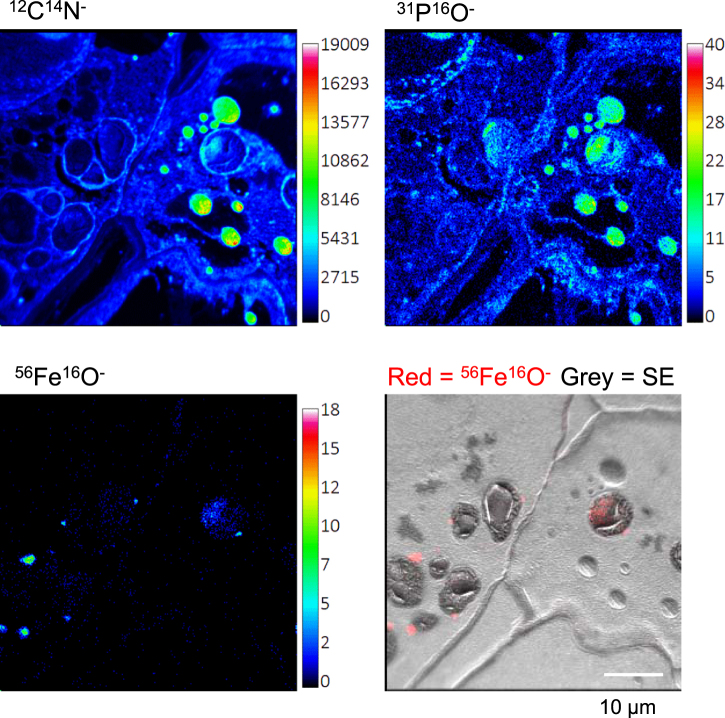
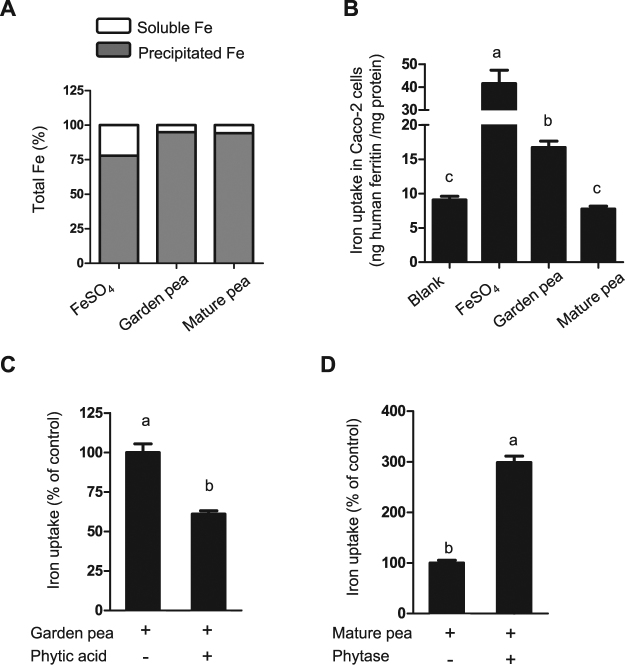
Pea seeds are widely consumed in their immature form, known as garden peas and petit pois, mostly after preservation by freezing or canning. Mature dry peas are rich in iron in the form of ferritin, but little is known about the content, form or bioavailability of iron in immature stages of seed development. Using specific antibodies and in-gel iron staining, we show that ferritin loaded with iron accumulated gradually during seed development. Immunolocalization and high-resolution secondary ion mass spectrometry (NanoSIMS) revealed that iron-loaded ferritin was located at the surface of starch-containing plastids. Standard cooking procedures destabilized monomeric ferritin and the iron-loaded form. Iron uptake studies using Caco-2 cells showed that the iron in microwaved immature peas was more bioavailable than in boiled mature peas, despite similar levels of soluble iron in the digestates. By manipulating the levels of phytic acid in the digestates we demonstrate that phytic acid is the main inhibitor of iron uptake from mature peas in vitro. Taken together, our data show that immature peas and mature dry peas contain similar levels of ferritin-iron, which is destabilized during cooking. However, iron from immature peas is more bioavailable because of lower phytic acid levels compared to mature peas.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
- BB/L025396/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/L025515/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/L013711/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/J004561/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/P012523/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
