

Rapid Phenotypic and Genotypic Diversification After Exposure to the Oral Host Niche in Candida albicans
- PMID: 29724862
- PMCID: PMC6028260
- DOI: 10.1534/genetics.118.301019
Rapid Phenotypic and Genotypic Diversification After Exposure to the Oral Host Niche in Candida albicans
Abstract
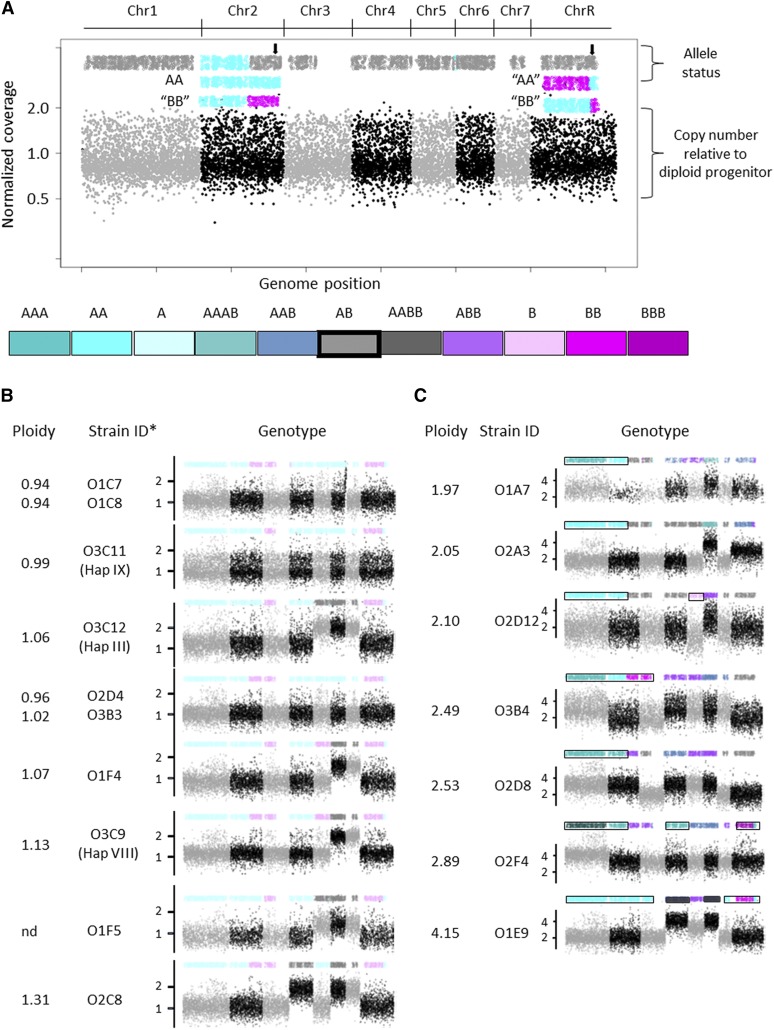
In vitro studies suggest that stress may generate random standing variation and that different cellular and ploidy states may evolve more rapidly under stress. Yet this idea has not been tested with pathogenic fungi growing within their host niche in vivo Here, we analyzed the generation of both genotypic and phenotypic diversity during exposure of Candida albicans to the mouse oral cavity. Ploidy, aneuploidy, loss of heterozygosity (LOH), and recombination were determined using flow cytometry and double digest restriction site-associated DNA sequencing. Colony phenotypic changes in size and filamentous growth were evident without selection and were enriched among colonies selected for LOH of the GAL1 marker. Aneuploidy and LOH occurred on all chromosomes (Chrs), with aneuploidy more frequent for smaller Chrs and whole Chr LOH more frequent for larger Chrs. Large genome shifts in ploidy to haploidy often maintained one or more heterozygous disomic Chrs, consistent with random Chr missegregation events. Most isolates displayed several different types of genomic changes, suggesting that the oral environment rapidly generates diversity de novo In sharp contrast, following in vitro propagation, isolates were not enriched for multiple LOH events, except in those that underwent haploidization and/or had high levels of Chr loss. The frequency of events was overall 100 times higher for C. albicans populations following in vivo passage compared with in vitro These hyper-diverse in vivo isolates likely provide C. albicans with the ability to adapt rapidly to the diversity of stress environments it encounters inside the host.
Keywords: Candida albicans; aneuploidy; colony phenotype; hyper-variability; loss of heterozygosity; oropharyngeal candidiasis.
Copyright © 2018 by the Genetics Society of America.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
