

MicroRNA-146a controls functional plasticity in γδ T cells by targeting NOD1
- PMID: 29728425
- PMCID: PMC7233287
- DOI: 10.1126/sciimmunol.aao1392
MicroRNA-146a controls functional plasticity in γδ T cells by targeting NOD1
Abstract
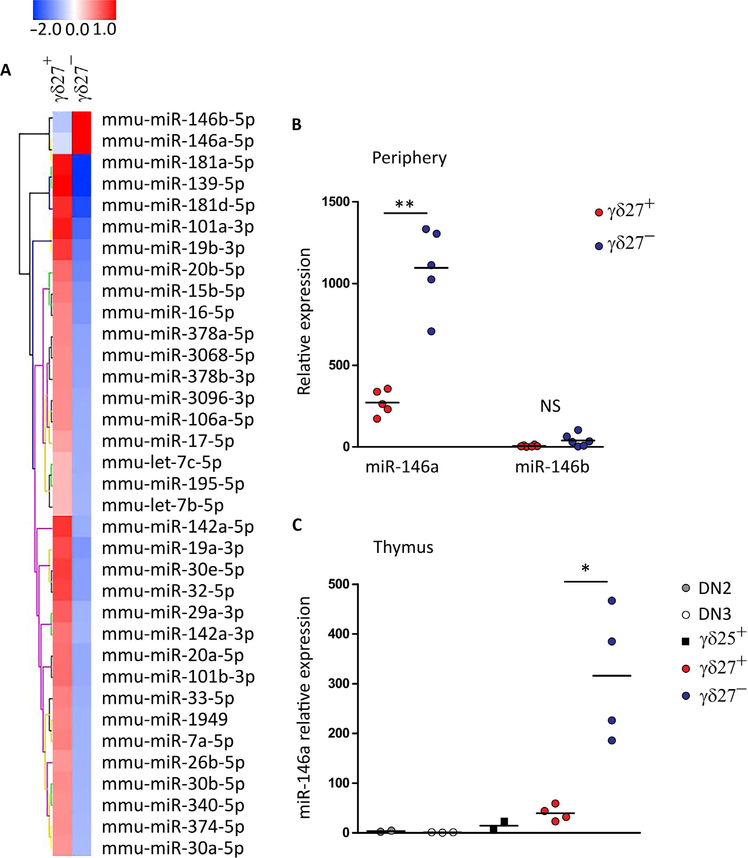
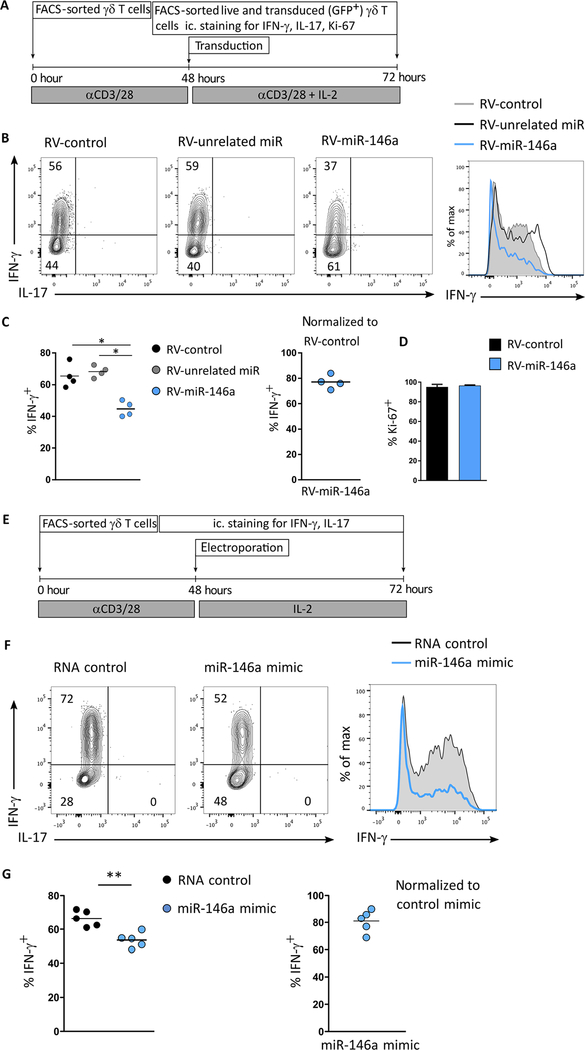
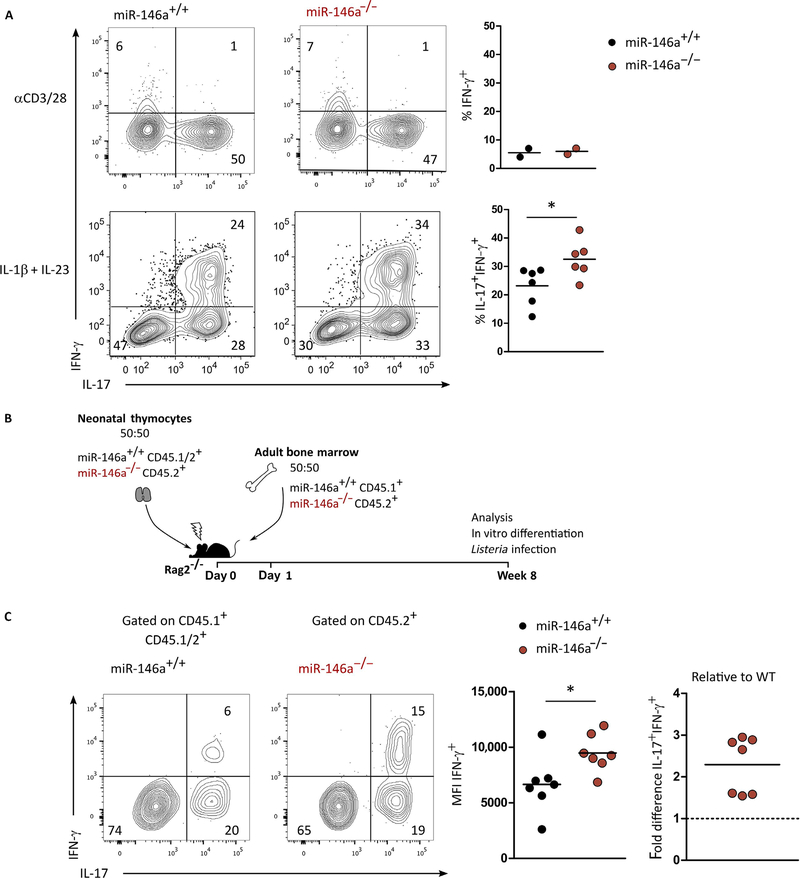
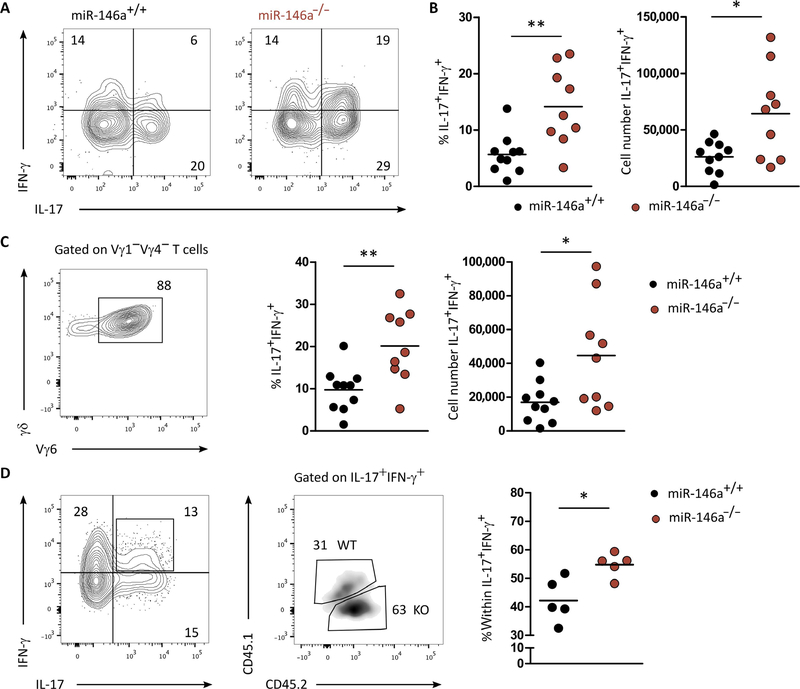
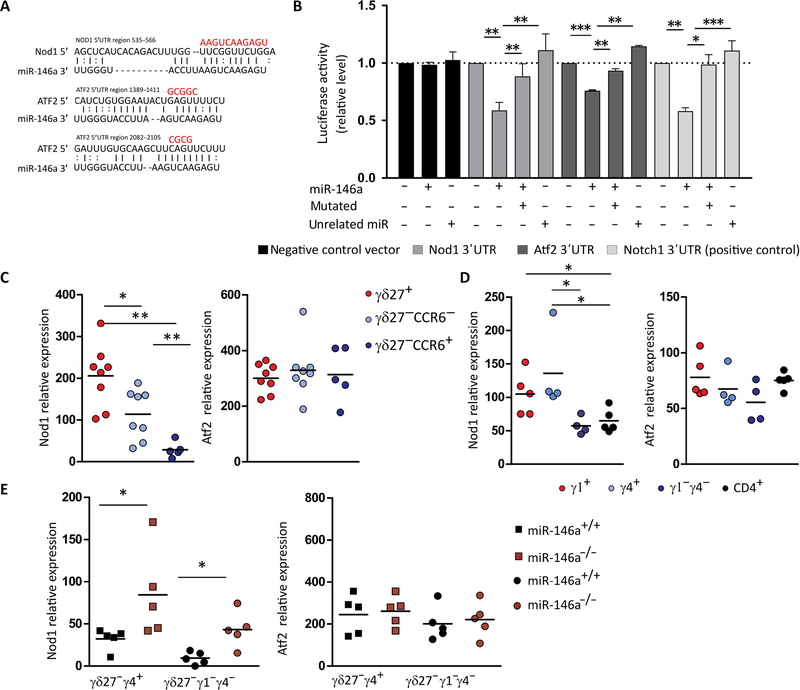
γδ T cells are major providers of proinflammatory cytokines. They are preprogrammed in the mouse thymus into distinct subsets producing either interleukin-17 (IL-17) or interferon-γ (IFN-γ), which segregate with CD27 expression. In the periphery, CD27- γδ (γδ27-) T cells can be induced under inflammatory conditions to coexpress IL-17 and IFN-γ; the molecular basis of this functional plasticity remains to be determined. On the basis of differential microRNA (miRNA) expression analysis and modulation in γδ T cell subsets, we identified miR-146a as a thymically imprinted post-transcriptional brake to limit IFN-γ expression in γδ27- T cells in vitro and in vivo. On the basis of biochemical purification of Argonaute 2-bound miR-146a targets, we identified Nod1 to be a relevant mRNA target that regulates γδ T cell plasticity. In line with this, Nod1-deficient mice lacked multifunctional IL-17+ IFN-γ+ γδ27- cells and were more susceptible to Listeria monocytogenes infection. Our studies establish the miR-146a/NOD1 axis as a key determinant of γδ T cell effector functions and plasticity.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures

References
-
- Korn T, Bettelli E, Oukka M, Kuchroo VK, IL-17 and Th17 Cells. Annu. Rev. Immunol. 27, 485–517 (2009). - PubMed
-
- Neurath MF, Weigmann B, Finotto S, Glickman J, Nieuwenhuis E, Iijima H, Mizoguchi A, Mizoguchi E, Mudter J, Galle PR, Bhan A, Autschbach F, Sullivan BM, Szabo SJ, Glimcher LH, Blumberg RS, The transcription factor T-bet regulates mucosal T cell activation in experimental colitis and Crohn’s disease. J. Exp. Med. 195, 1129–1143 (2002). - PMC - PubMed
-
- Papotto PH, Ribot JC, Silva-Santos B, IL-17+ γδ T cells as kick-starters of inflammation. Nat. Immunol. 18, 604–611 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
