

The flagellar protein Enkurin is required for mouse sperm motility and for transport through the female reproductive tract
- PMID: 29733335
- PMCID: PMC6203878
- DOI: 10.1093/biolre/ioy105
The flagellar protein Enkurin is required for mouse sperm motility and for transport through the female reproductive tract
Abstract
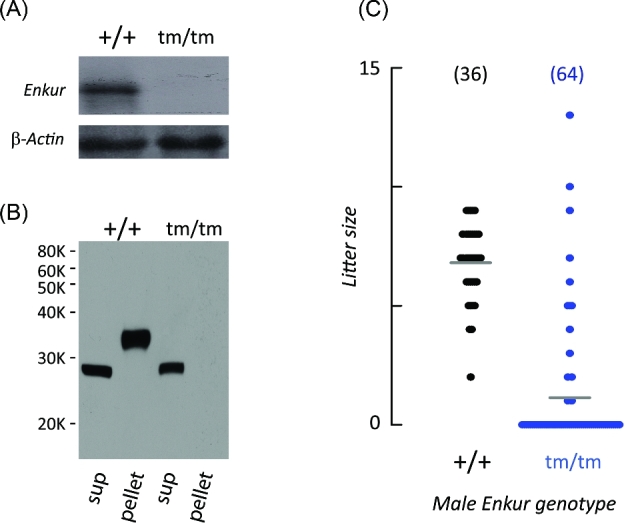
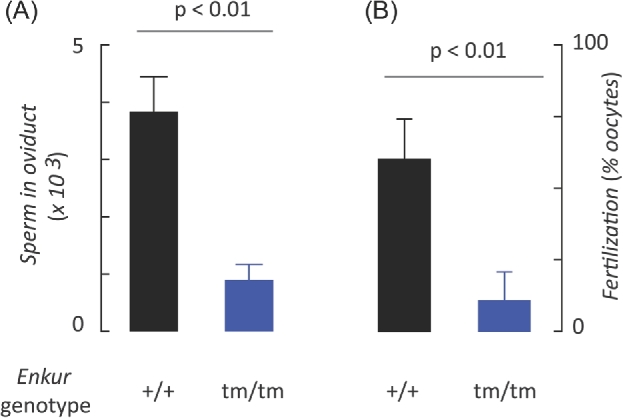
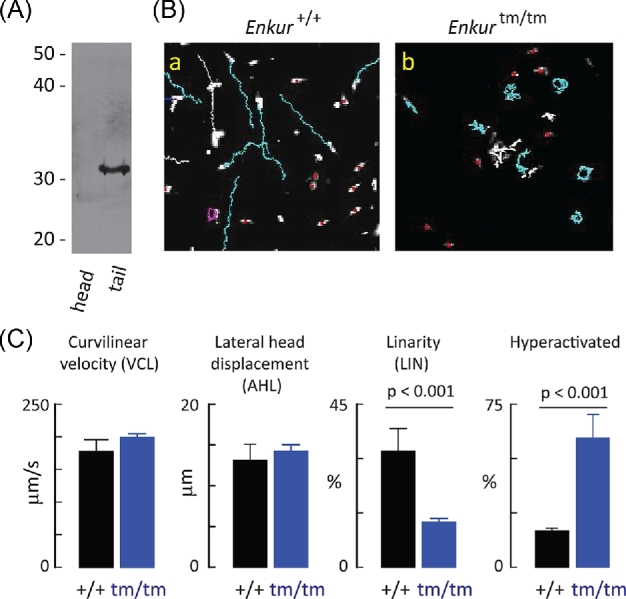
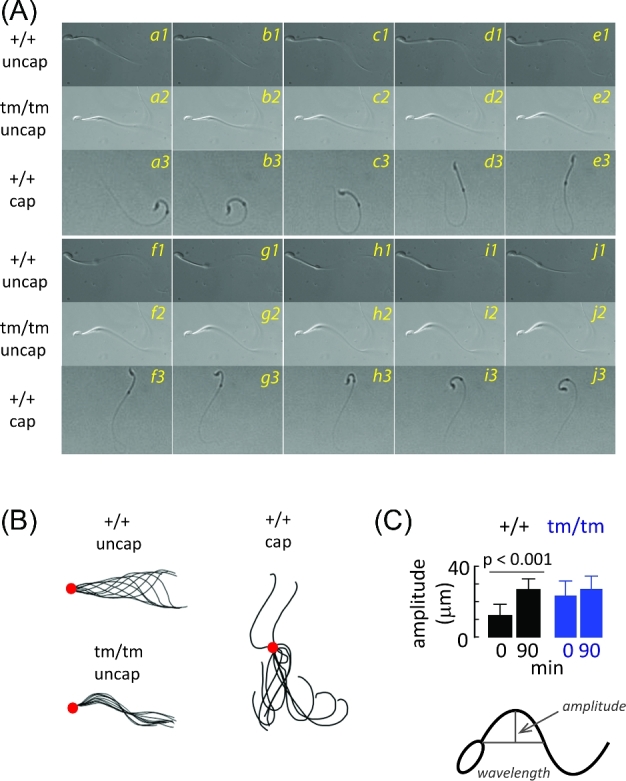
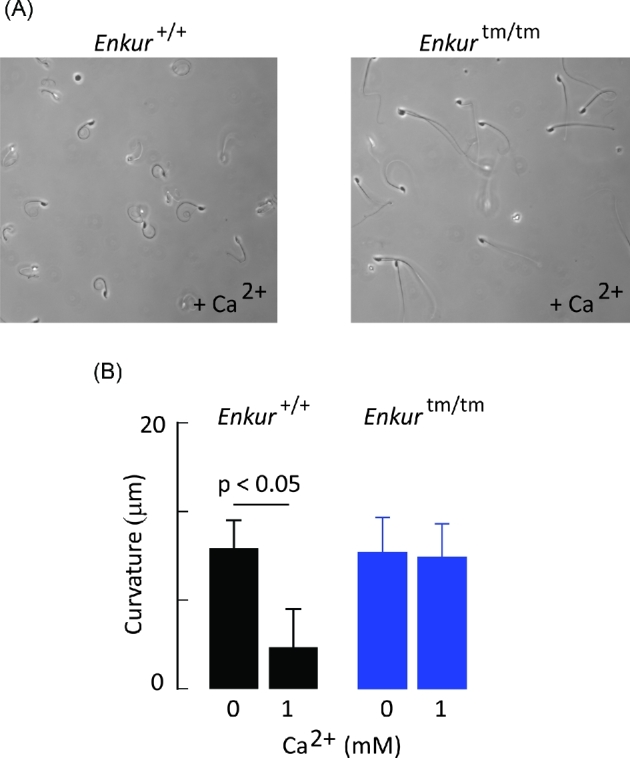
Enkurin was identified initially in mouse sperm where it was suggested to act as an intracellular adaptor protein linking membrane calcium influx to intracellular signaling pathways. In order to examine the function of this protein, a targeted mutation was introduced into the mouse Enkurin gene. Males that were homozygous for this mutated allele were subfertile. This was associated with lower rates of sperm transport in the female reproductive tract, including reduced entry into the oviduct and slower migration to the site of fertilization in the distal oviduct, and with poor progressive motility in vitro. Flagella from wild-type animals exhibited symmetrical bending and progressive motility in culture medium, and demembranated flagella exhibited the "curlicue" response to Ca2+ in vitro. In contrast, flagella of mice homozygous for the mutated allele displayed only asymmetric bending, nonprogressive motility, and a loss of Ca2+-responsiveness following demembrantion. We propose that Enkurin is part of a flagellar Ca2+-sensor that regulates bending and that the motility defects following mutation of the locus are the proximate cause of subfertility.
Figures
Similar articles
-
Enkurin is a novel calmodulin and TRPC channel binding protein in sperm.Dev Biol. 2004 Oct 15;274(2):426-35. doi: 10.1016/j.ydbio.2004.07.031. Dev Biol. 2004. PMID: 15385169
-
Functional deficiencies and a reduced response to calcium in the flagellum of mouse sperm lacking SPAG16L.Biol Reprod. 2010 Apr;82(4):736-44. doi: 10.1095/biolreprod.109.080143. Epub 2009 Dec 30. Biol Reprod. 2010. PMID: 20042536 Free PMC article.
-
Mapping and cloning recombinant breakpoints demarcating the hybrid sterility 6-specific sperm tail assembly defect.Mamm Genome. 1999 Feb;10(2):88-94. doi: 10.1007/s003359900950. Mamm Genome. 1999. PMID: 9922385
-
Tales from the tail: what do we really know about sperm motility?J Androl. 2003 Nov-Dec;24(6):790-803. doi: 10.1002/j.1939-4640.2003.tb03123.x. J Androl. 2003. PMID: 14581499 Review. No abstract available.
-
Factors controlling sperm migration through the oviduct revealed by gene-modified mouse models.Exp Anim. 2018 May 10;67(2):91-104. doi: 10.1538/expanim.17-0153. Epub 2018 Jan 22. Exp Anim. 2018. PMID: 29353867 Free PMC article. Review.
Cited by
-
Enkurin: A novel marker for myeloproliferative neoplasms from platelet, megakaryocyte, and whole blood specimens.bioRxiv [Preprint]. 2023 Jan 12:2023.01.07.523111. doi: 10.1101/2023.01.07.523111. bioRxiv. 2023. Update in: Blood Adv. 2023 Sep 26;7(18):5433-5445. doi: 10.1182/bloodadvances.2022008939. PMID: 36712071 Free PMC article. Updated. Preprint.
-
Evolutionary adaptations of doublet microtubules in trypanosomatid parasites.Science. 2025 Mar 14;387(6739):eadr5507. doi: 10.1126/science.adr5507. Epub 2025 Mar 14. Science. 2025. PMID: 40080577 Free PMC article.
-
De novo identification of mammalian ciliary motility proteins using cryo-EM.Cell. 2021 Nov 11;184(23):5791-5806.e19. doi: 10.1016/j.cell.2021.10.007. Epub 2021 Oct 28. Cell. 2021. PMID: 34715025 Free PMC article.
-
CFAP45 deficiency causes situs abnormalities and asthenospermia by disrupting an axonemal adenine nucleotide homeostasis module.Nat Commun. 2020 Nov 2;11(1):5520. doi: 10.1038/s41467-020-19113-0. Nat Commun. 2020. PMID: 33139725 Free PMC article.
-
The inner junction protein CFAP20 functions in motile and non-motile cilia and is critical for vision.Nat Commun. 2022 Nov 3;13(1):6595. doi: 10.1038/s41467-022-33820-w. Nat Commun. 2022. PMID: 36329026 Free PMC article.
References
-
- Sutton KA, Jungnickel MK, Wang Y, Cullen K, Lambert S, Florman HM. Enkurin is a novel calmodulin and TRPC channel binding protein in sperm. Dev Biol 2004; 274(2):426–435. - PubMed
-
- Nakachi M, Nakajima A, Nomura M, Yonezawa K, Ueno K, Endo T, Inaba K. Proteomic profiling reveals compartment-specific, novel functions of ascidian sperm proteins. Mol Reprod Dev 2011; 78:529–549. - PubMed
-
- Bartel M, Hartmann S, Lehmann K, Postel K, Quesada H, Philipp EER, Heilmann K, Micheel B, Stuckas H. Identification of sperm proteins as candidate biomarkers for the analysis of reproductive isolation in Mytilus: a case study for the enkurin locus. Mar Biol 2012; 159(10):2195–2207.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
