

PII-like signaling protein SbtB links cAMP sensing with cyanobacterial inorganic carbon response
- PMID: 29735650
- PMCID: PMC6003466
- DOI: 10.1073/pnas.1803790115
PII-like signaling protein SbtB links cAMP sensing with cyanobacterial inorganic carbon response
Abstract
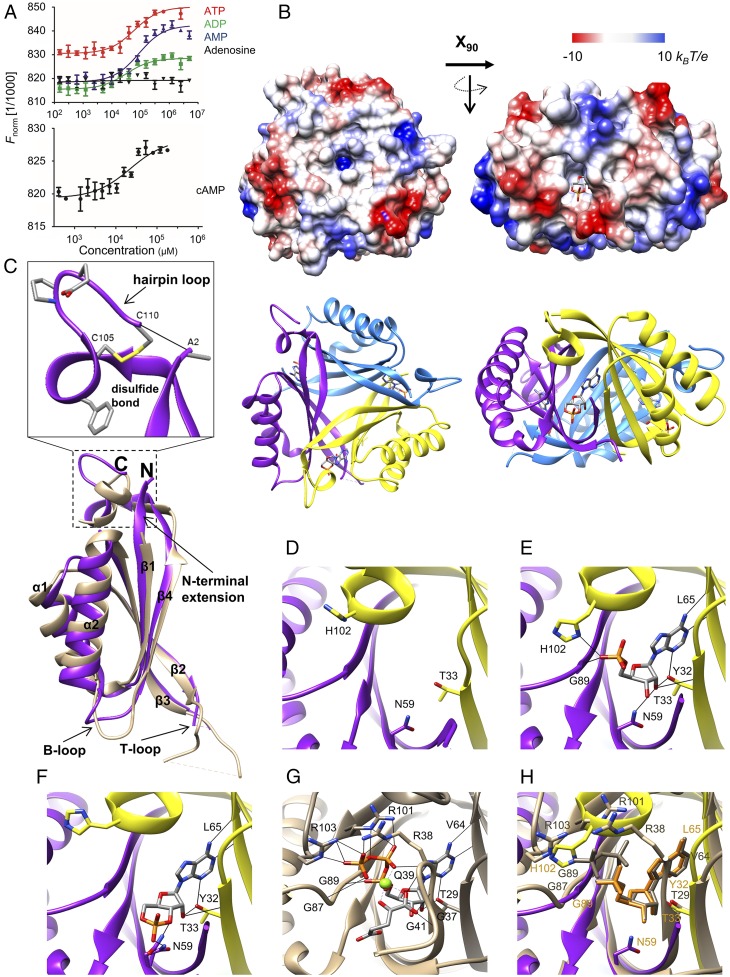
Cyanobacteria are phototrophic prokaryotes that evolved oxygenic photosynthesis ∼2.7 billion y ago and are presently responsible for ∼10% of total global photosynthetic production. To cope with the evolutionary pressure of dropping ambient CO2 concentrations, they evolved a CO2-concentrating mechanism (CCM) to augment intracellular inorganic carbon (Ci) levels for efficient CO2 fixation. However, how cyanobacteria sense the fluctuation in Ci is poorly understood. Here we present biochemical, structural, and physiological insights into SbtB, a unique PII-like signaling protein, which provides new insights into Ci sensing. SbtB is highly conserved in cyanobacteria and is coexpressed with CCM genes. The SbtB protein from the cyanobacterium Synechocystis sp. PCC 6803 bound a variety of adenosine nucleotides, including the second messenger cAMP. Cocrystal structures unraveled the individual binding modes of trimeric SbtB with AMP and cAMP. The nucleotide-binding pocket is located between the subunit clefts of SbtB, perfectly matching the structure of canonical PII proteins. This clearly indicates that proteins of the PII superfamily arose from a common ancestor, whose structurally conserved nucleotide-binding pocket has evolved to sense different adenyl nucleotides for various signaling functions. Moreover, we provide physiological and biochemical evidence for the involvement of SbtB in Ci acclimation. Collectively, our results suggest that SbtB acts as a Ci sensor protein via cAMP binding, highlighting an evolutionarily conserved role for cAMP in signaling the cellular carbon status.
Keywords: PII-like protein SbtB; cAMP; cyanobacteria; inorganic carbon signaling; signal transduction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Carbon/nitrogen homeostasis control in cyanobacteria.FEMS Microbiol Rev. 2020 Jan 1;44(1):33-53. doi: 10.1093/femsre/fuz025. FEMS Microbiol Rev. 2020. PMID: 31617886 Free PMC article. Review.
-
The redox-sensitive R-loop of the carbon control protein SbtB contributes to the regulation of the cyanobacterial CCM.Sci Rep. 2024 Apr 3;14(1):7885. doi: 10.1038/s41598-024-58354-7. Sci Rep. 2024. PMID: 38570698 Free PMC article.
-
Carbon signaling protein SbtB possesses atypical redox-regulated apyrase activity to facilitate regulation of bicarbonate transporter SbtA.Proc Natl Acad Sci U S A. 2023 Feb 21;120(8):e2205882120. doi: 10.1073/pnas.2205882120. Epub 2023 Feb 17. Proc Natl Acad Sci U S A. 2023. PMID: 36800386 Free PMC article.
-
The impact of the cyanobacterial carbon-regulator protein SbtB and of the second messengers cAMP and c-di-AMP on CO2 -dependent gene expression.New Phytol. 2022 Jun;234(5):1801-1816. doi: 10.1111/nph.18094. Epub 2022 Apr 9. New Phytol. 2022. PMID: 35285042
-
Roles of second messengers in the regulation of cyanobacterial physiology: the carbon-concentrating mechanism and beyond.Microlife. 2023 Feb 23;4:uqad008. doi: 10.1093/femsml/uqad008. eCollection 2023. Microlife. 2023. PMID: 37223741 Free PMC article. Review.
Cited by
-
Tetragonal crystal form of the cyanobacterial bicarbonate-transporter regulator SbtB from Synechocystis sp. PCC 6803.Acta Crystallogr F Struct Biol Commun. 2020 Sep 1;76(Pt 9):438-443. doi: 10.1107/S2053230X20010523. Epub 2020 Aug 19. Acta Crystallogr F Struct Biol Commun. 2020. PMID: 32880592 Free PMC article.
-
Bacterial Adaptation by a Transposition Burst of an Invading IS Element.Genome Biol Evol. 2021 Nov 5;13(11):evab245. doi: 10.1093/gbe/evab245. Genome Biol Evol. 2021. PMID: 34791212 Free PMC article.
-
The second messenger c-di-AMP controls natural competence via ComFB signaling protein.Cell Discov. 2025 Aug 1;11(1):65. doi: 10.1038/s41421-025-00816-x. Cell Discov. 2025. PMID: 40745163 Free PMC article. No abstract available.
-
Carbon/nitrogen homeostasis control in cyanobacteria.FEMS Microbiol Rev. 2020 Jan 1;44(1):33-53. doi: 10.1093/femsre/fuz025. FEMS Microbiol Rev. 2020. PMID: 31617886 Free PMC article. Review.
-
The redox-sensitive R-loop of the carbon control protein SbtB contributes to the regulation of the cyanobacterial CCM.Sci Rep. 2024 Apr 3;14(1):7885. doi: 10.1038/s41598-024-58354-7. Sci Rep. 2024. PMID: 38570698 Free PMC article.
References
-
- Hohmann-Marriott MF, Blankenship RE. Evolution of photosynthesis. Annu Rev Plant Biol. 2011;62:515–548. - PubMed
-
- Shibata M, et al. Genes essential to sodium-dependent bicarbonate transport in cyanobacteria: Function and phylogenetic analysis. J Biol Chem. 2002;277:18658–18664. - PubMed
-
- Wang HL, Postier BL, Burnap RL. Alterations in global patterns of gene expression in Synechocystis sp. PCC 6803 in response to inorganic carbon limitation and the inactivation of ndhR, a LysR family regulator. J Biol Chem. 2004;279:5739–5751. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
