

Defective phagosome motility and degradation in cell nonautonomous RPE pathogenesis of a dominant macular degeneration
- PMID: 29735674
- PMCID: PMC6003516
- DOI: 10.1073/pnas.1709211115
Defective phagosome motility and degradation in cell nonautonomous RPE pathogenesis of a dominant macular degeneration
Abstract
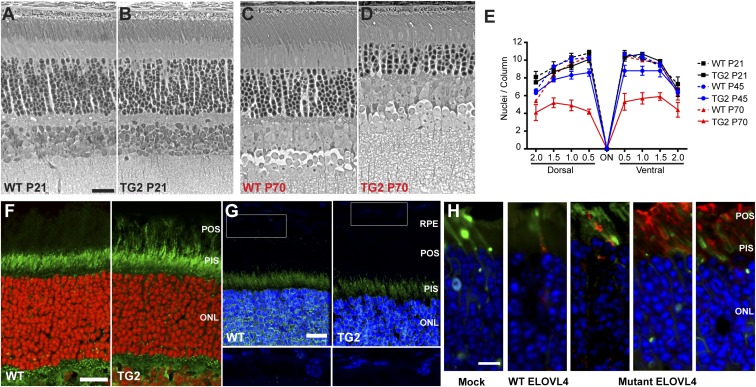
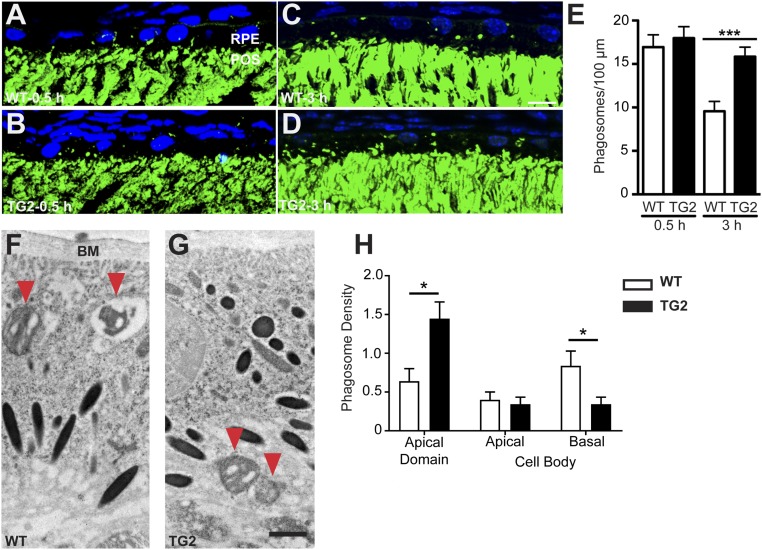
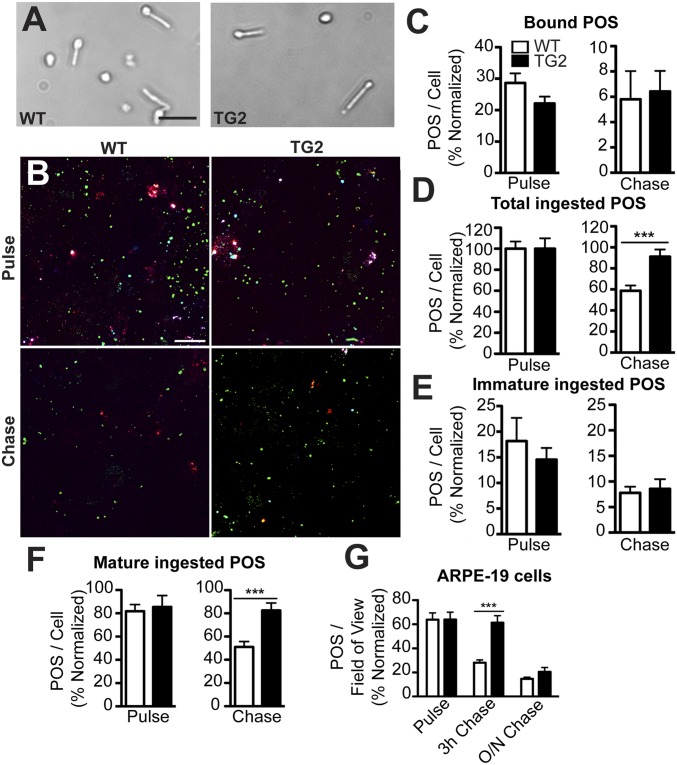
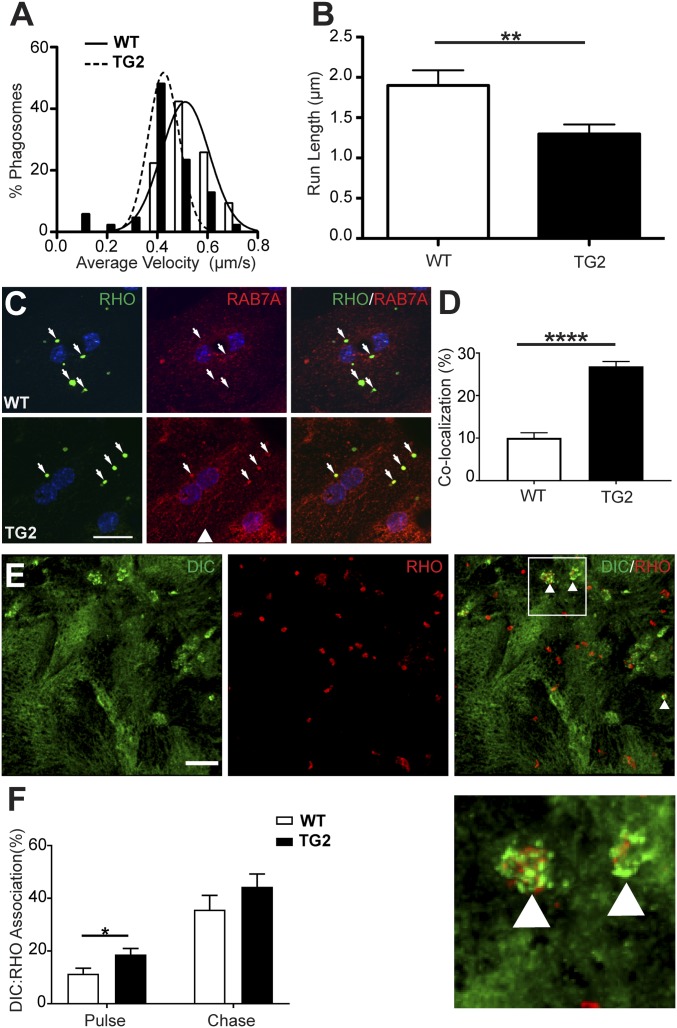
Stargardt macular dystrophy 3 (STGD3) is caused by dominant mutations in the ELOVL4 gene. Like other macular degenerations, pathogenesis within the retinal pigment epithelium (RPE) appears to contribute to the loss of photoreceptors from the central retina. However, the RPE does not express ELOVL4, suggesting photoreceptor cell loss in STGD3 occurs through two cell nonautonomous events: mutant photoreceptors first affect RPE cell pathogenesis, and then, second, RPE dysfunction leads to photoreceptor cell death. Here, we have investigated how the RPE pathology occurs, using a STGD3 mouse model in which mutant human ELOVL4 is expressed in the photoreceptors. We found that the mutant protein was aberrantly localized to the photoreceptor outer segment (POS), and that resulting POS phagosomes were degraded more slowly in the RPE. In cell culture, the mutant POSs are ingested by primary RPE cells normally, but the phagosomes are processed inefficiently, even by wild-type RPE. The mutant phagosomes excessively sequester RAB7A and dynein, and have impaired motility. We propose that the abnormal presence of ELOVL4 protein in POSs results in phagosomes that are defective in recruiting appropriate motor protein linkers, thus contributing to slower degradation because their altered motility results in slower basal migration and fewer productive encounters with endolysosomes. In the transgenic mouse retinas, the RPE accumulated abnormal-looking phagosomes and oxidative stress adducts; these pathological changes were followed by pathology in the neural retina. Our results indicate inefficient phagosome degradation as a key component of the first cell nonautonomous event underlying retinal degeneration due to mutant ELOVL4.
Keywords: ELOVL4; Stargardt; phagocytosis; photoreceptor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Zhang K, et al. A 5-bp deletion in ELOVL4 is associated with two related forms of autosomal dominant macular dystrophy. Nat Genet. 2001;27:89–93. - PubMed
-
- Edwards AO, Donoso LA, Ritter R., 3rd A novel gene for autosomal dominant Stargardt-like macular dystrophy with homology to the SUR4 protein family. Invest Ophthalmol Vis Sci. 2001;42:2652–2663. - PubMed
-
- Bernstein PS, et al. Diverse macular dystrophy phenotype caused by a novel complex mutation in the ELOVL4 gene. Invest Ophthalmol Vis Sci. 2001;42:3331–3336. - PubMed
-
- Maugeri A, et al. A novel mutation in the ELOVL4 gene causes autosomal dominant Stargardt-like macular dystrophy. Invest Ophthalmol Vis Sci. 2004;45:4263–4267. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
