

A non-equilibrium approach to allosteric communication
- PMID: 29735740
- PMCID: PMC5941181
- DOI: 10.1098/rstb.2017.0187
A non-equilibrium approach to allosteric communication
Abstract
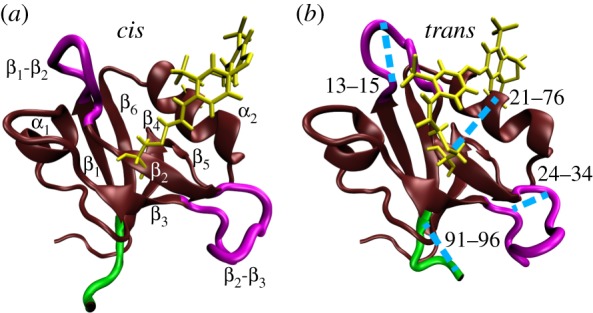
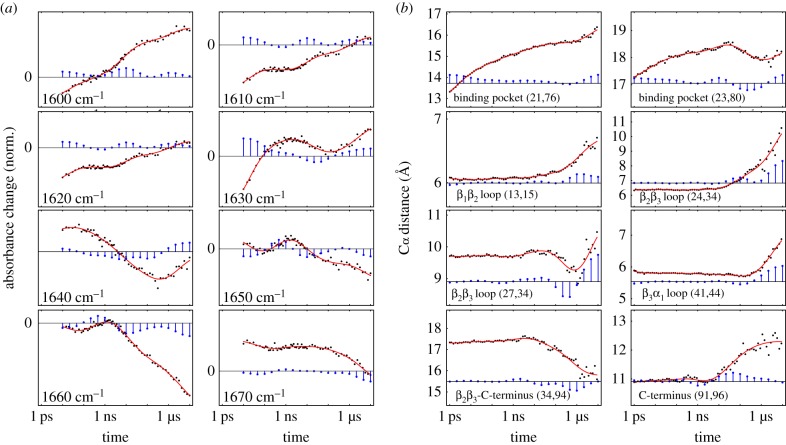
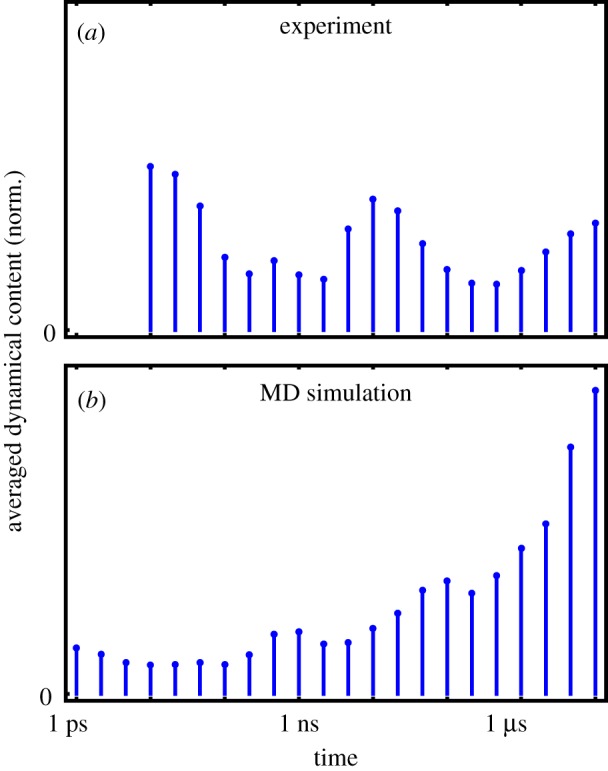
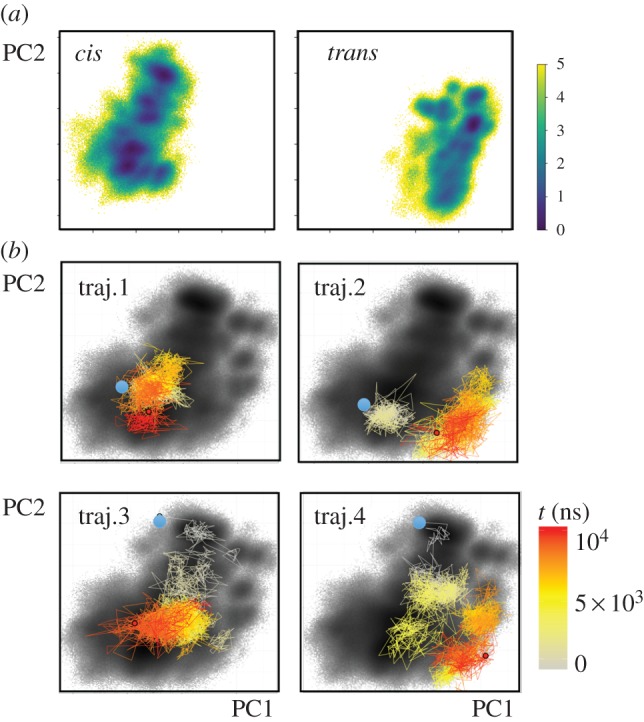
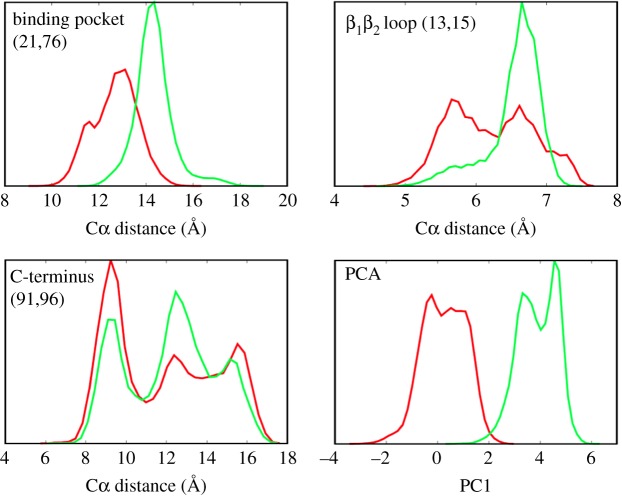
While the theory of protein folding is well developed, including concepts such as rugged energy landscape, folding funnel, etc., the same degree of understanding has not been reached for the description of the dynamics of allosteric transitions in proteins. This is not only due to the small size of the structural change upon ligand binding to an allosteric site, but also due to challenges in designing experiments that directly observe such an allosteric transition. On the basis of recent pump-probe-type experiments (Buchli et al. 2013 Proc. Natl Acad. Sci. USA110, 11 725-11 730. (doi:10.1073/pnas.1306323110)) and non-equilibrium molecular dynamics simulations (Buchenberg et al. 2017 Proc. Natl Acad. Sci. USA114, E6804-E6811. (doi:10.1073/pnas.1707694114)) studying an photoswitchable PDZ2 domain as model for an allosteric transition, we outline in this perspective how such a description of allosteric communication might look. That is, calculating the dynamical content of both experiment and simulation (which agree remarkably well with each other), we find that allosteric communication shares some properties with downhill folding, except that it is an 'order-order' transition. Discussing the multiscale and hierarchical features of the dynamics, the validity of linear response theory as well as the meaning of 'allosteric pathways', we conclude that non-equilibrium experiments and simulations are a promising way to study dynamical aspects of allostery.This article is part of a discussion meeting issue 'Allostery and molecular machines'.
Keywords: allosteric transition; downhill folding; dynamic content; free-energy landscape; non-equilibrium molecular dynamics simulations; time-resolved vibrational spectroscopy.
© 2018 The Author(s).
Conflict of interest statement
We declare we have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
