

A High Frequency of HIV-Specific Circulating Follicular Helper T Cells Is Associated with Preserved Memory B Cell Responses in HIV Controllers
- PMID: 29739909
- PMCID: PMC5941072
- DOI: 10.1128/mBio.00317-18
A High Frequency of HIV-Specific Circulating Follicular Helper T Cells Is Associated with Preserved Memory B Cell Responses in HIV Controllers
Abstract
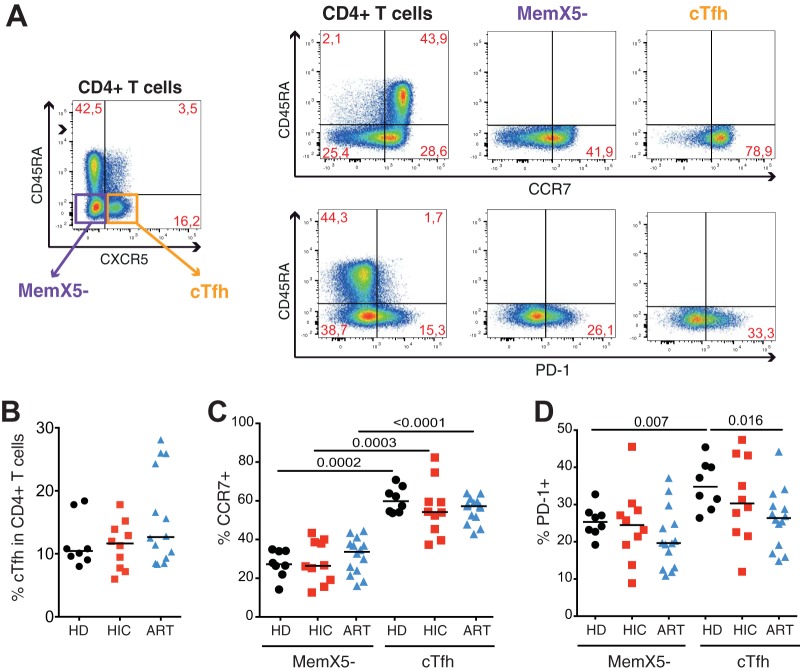
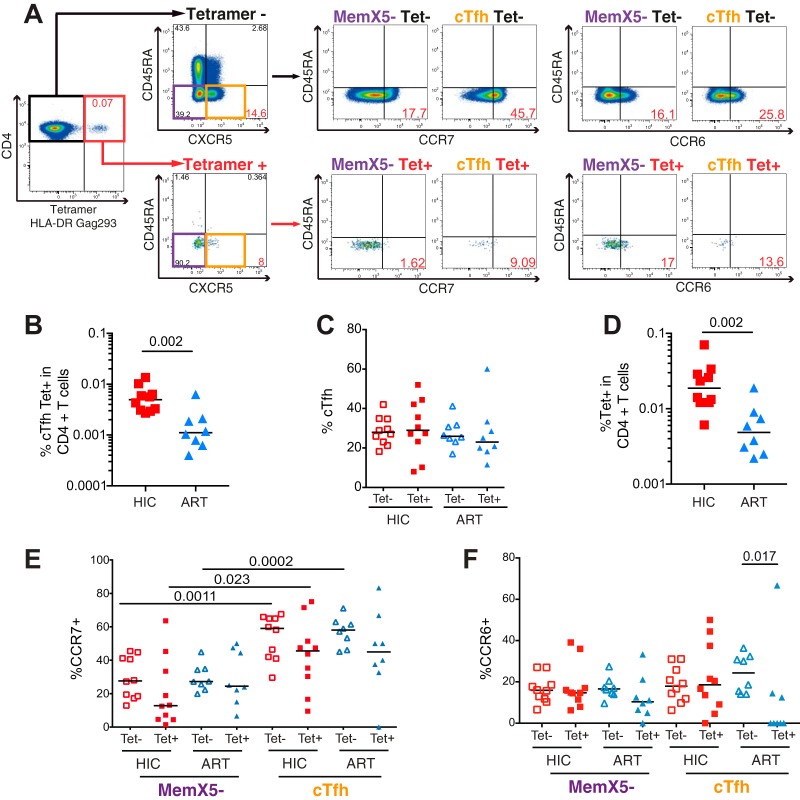
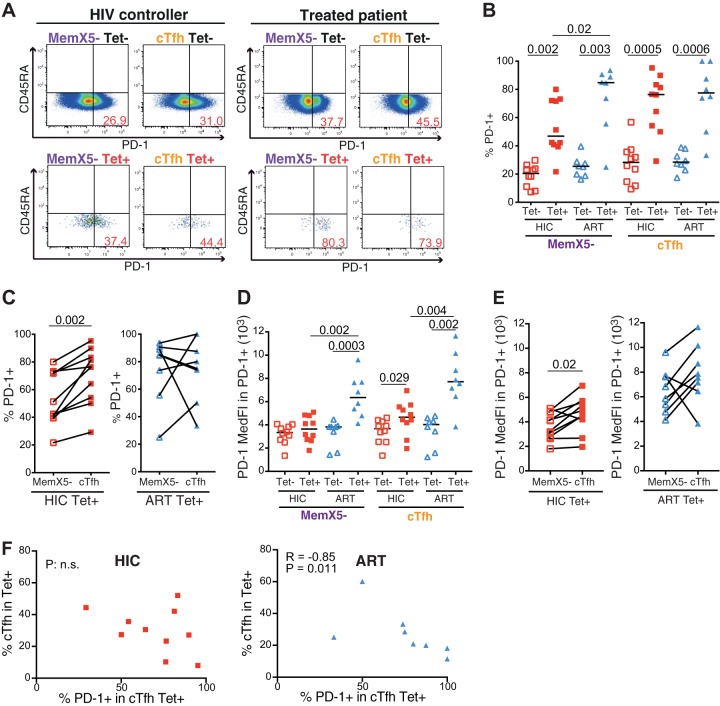
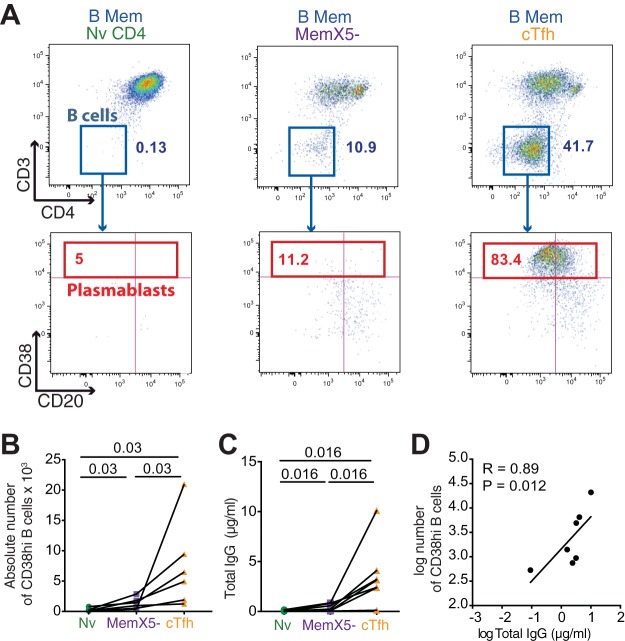
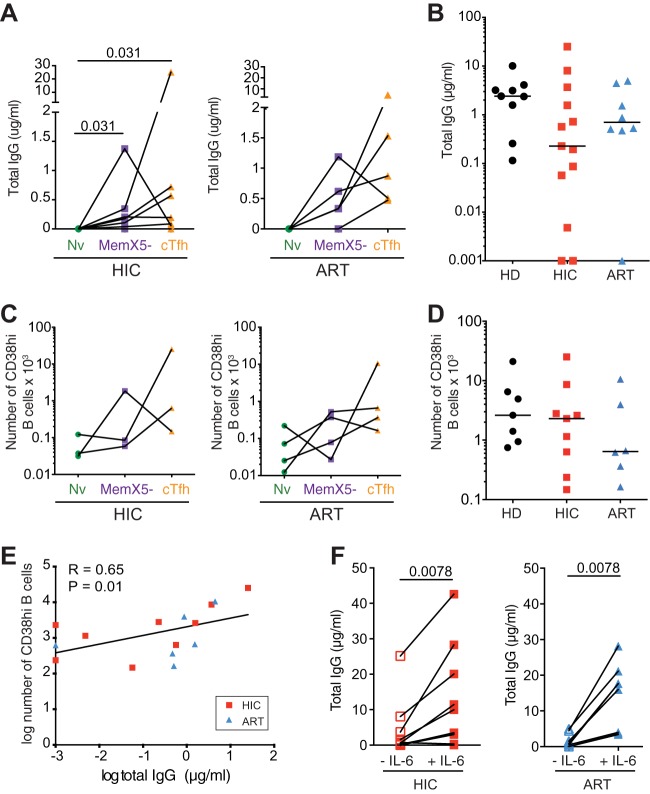
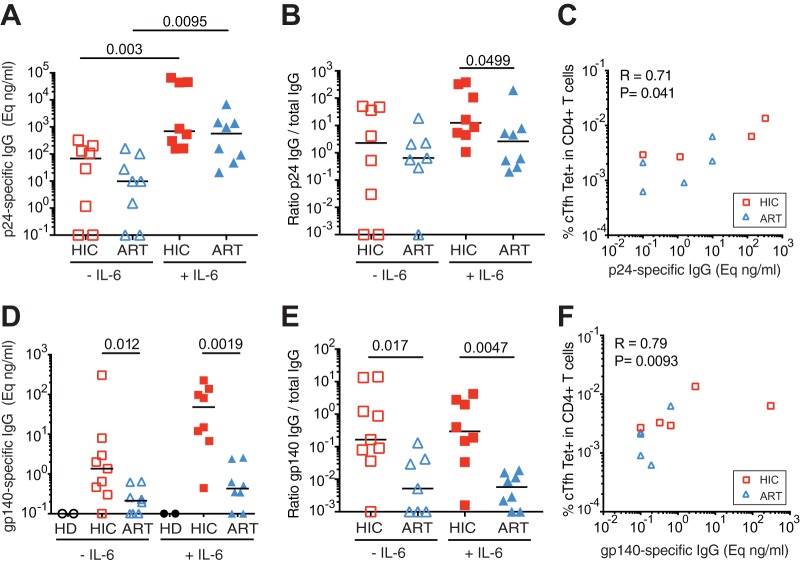
Follicular helper T cells (Tfh) play an essential role in the affinity maturation of the antibody response by providing help to B cells. To determine whether this CD4+ T cell subset may contribute to the spontaneous control of HIV infection, we analyzed the phenotype and function of circulating Tfh (cTfh) in patients from the ANRS CO21 CODEX cohort who naturally controlled HIV-1 replication to undetectable levels and compared them to treated patients with similarly low viral loads. HIV-specific cTfh (Tet+), detected by Gag-major histocompatibility complex class II (MHC-II) tetramer labeling in the CD45RA- CXCR5+ CD4+ T cell population, proved more frequent in the controller group (P = 0.002). The frequency of PD-1 expression in Tet+ cTfh was increased in both groups (median, >75%) compared to total cTfh (<30%), but the intensity of PD-1 expression per cell remained higher in the treated patient group (P = 0.02), pointing to the persistence of abnormal immune activation in treated patients. The function of cTfh, analyzed by the capacity to promote IgG secretion in cocultures with autologous memory B cells, did not show major differences between groups in terms of total IgG production but proved significantly more efficient in the controller group when measuring HIV-specific IgG production. The frequency of Tet+ cTfh correlated with HIV-specific IgG production (R = 0.71 for Gag-specific and R = 0.79 for Env-specific IgG, respectively). Taken together, our findings indicate that key cTfh-B cell interactions are preserved in controlled HIV infection, resulting in potent memory B cell responses that may play an underappreciated role in HIV control.IMPORTANCE The rare patients who spontaneously control HIV replication in the absence of therapy provide a unique model to identify determinants of an effective anti-HIV immune response. HIV controllers show signs of particularly efficient antiviral T cell responses, while their humoral response was until recently considered to play only a minor role in viral control. However, emerging evidence suggests that HIV controllers maintain a significant but "silent" antiviral memory B cell population that can be reactivated upon antigenic stimulation. We report that cTfh help likely contributes to the persistence of controller memory B cell responses, as the frequency of HIV-specific cTfh correlated with the induction of HIV-specific antibodies in functional assays. These findings suggest that T follicular help may contribute to HIV control and highlight the need for inducing such help in HIV vaccine strategies that aim at eliciting persistent B cell responses.
Keywords: B lymphocytes; HIV controllers; MHC-II tetramers; T follicular helper; T lymphocytes; human immunodeficiency virus.
Copyright © 2018 Claireaux et al.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
