

Systematic analysis of DEMETER-like DNA glycosylase genes shows lineage-specific Smi-miR7972 involved in SmDML1 regulation in Salvia miltiorrhiza
- PMID: 29739980
- PMCID: PMC5940787
- DOI: 10.1038/s41598-018-25315-w
Systematic analysis of DEMETER-like DNA glycosylase genes shows lineage-specific Smi-miR7972 involved in SmDML1 regulation in Salvia miltiorrhiza
Abstract
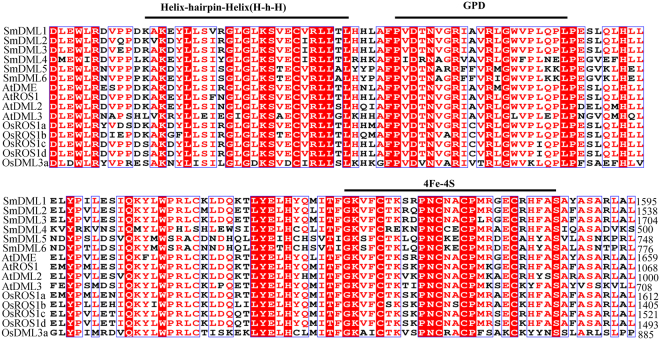
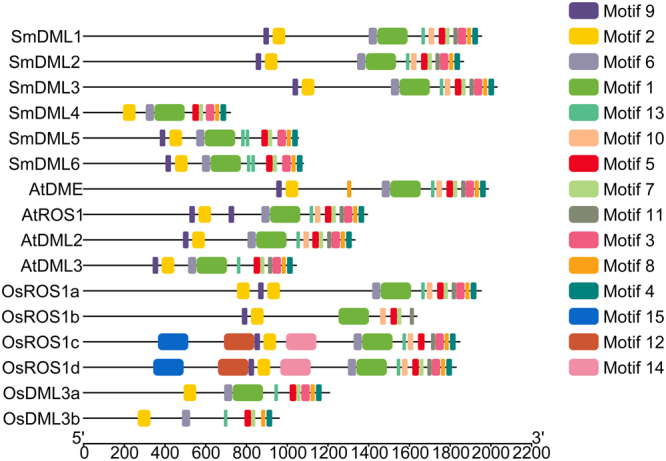
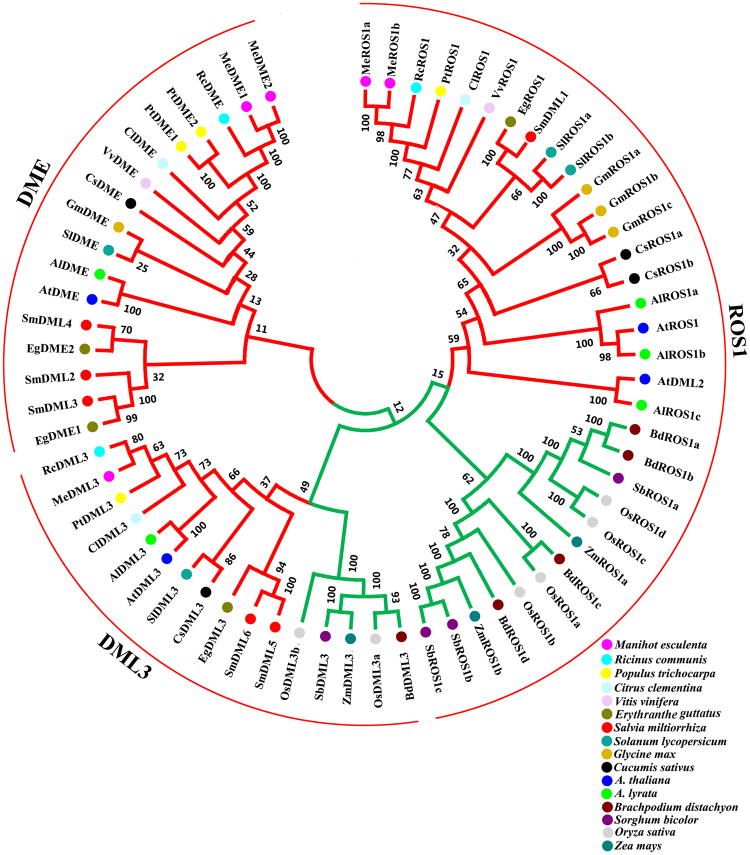
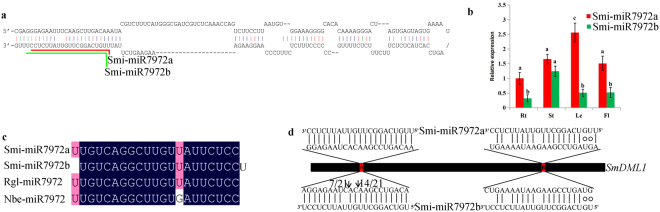
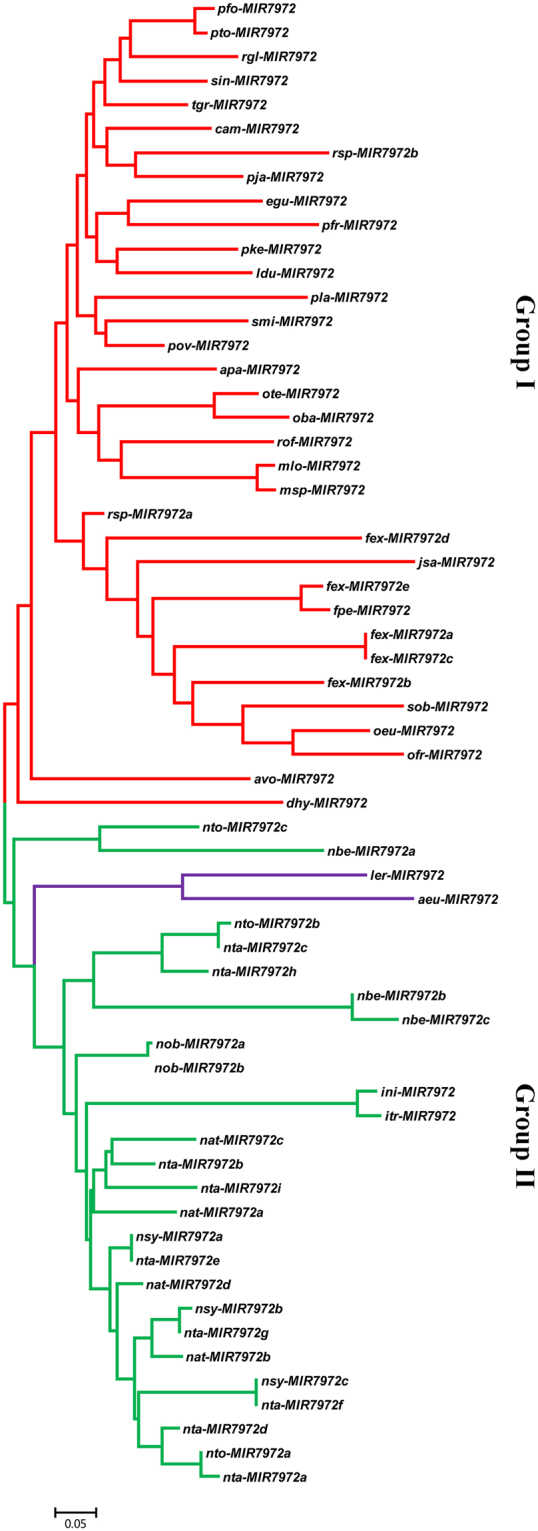
DEMETER-like DNA glycosylases (DMLs) initiate the base excision repair-dependent DNA demethylation to regulate a wide range of biological processes in plants. Six putative SmDML genes, termed SmDML1-SmDML6, were identified from the genome of S. miltiorrhiza, an emerging model plant for Traditional Chinese Medicine (TCM) studies. Integrated analysis of gene structures, sequence features, conserved domains and motifs, phylogenetic analysis and differential expression showed the conservation and divergence of SmDMLs. SmDML1, SmDML2 and SmDML4 were significantly down-regulated by the treatment of 5Aza-dC, a general DNA methylation inhibitor, suggesting involvement of SmDMLs in genome DNA methylation change. SmDML1 was predicted and experimentally validated to be target of Smi-miR7972. Computational analysis of forty whole genome sequences and almost all of RNA-seq data from Lamiids revealed that MIR7972s were only distributed in some plants of the three orders, including Lamiales, Solanales and Boraginales, and the number of MIR7972 genes varied among species. It suggests that MIR7972 genes underwent expansion and loss during the evolution of some Lamiids species. Phylogenetic analysis of MIR7972s showed closer evolutionary relationships between MIR7972s in Boraginales and Solanales in comparison with Lamiales. These results provide a valuable resource for elucidating DNA demethylation mechanism in S. miltiorrhiza.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
