

Breathing matters
- PMID: 29740175
- PMCID: PMC6636643
- DOI: 10.1038/s41583-018-0003-6
Breathing matters
Abstract
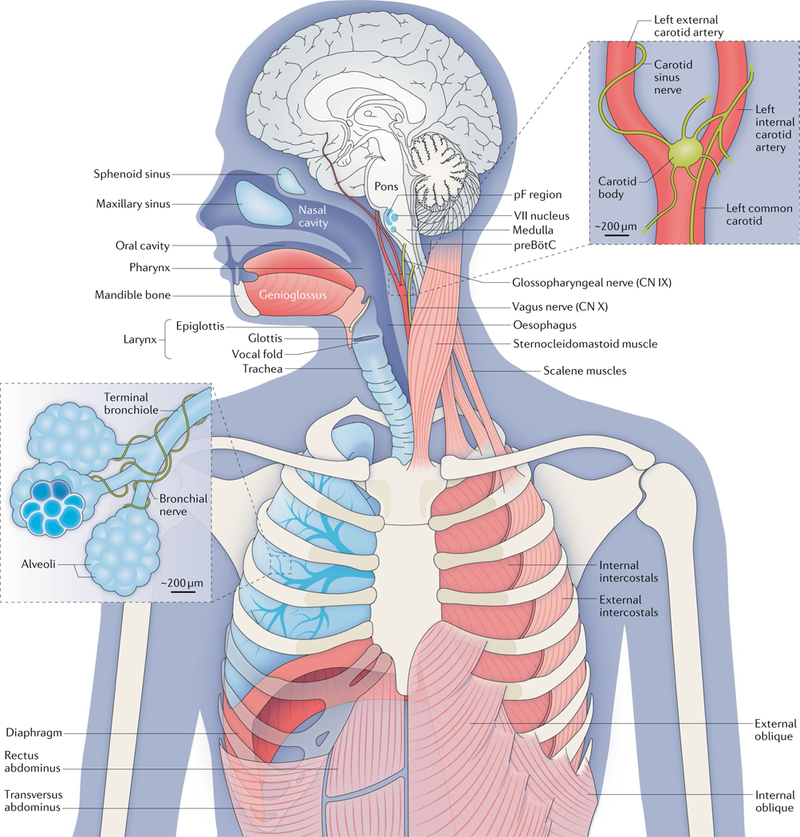
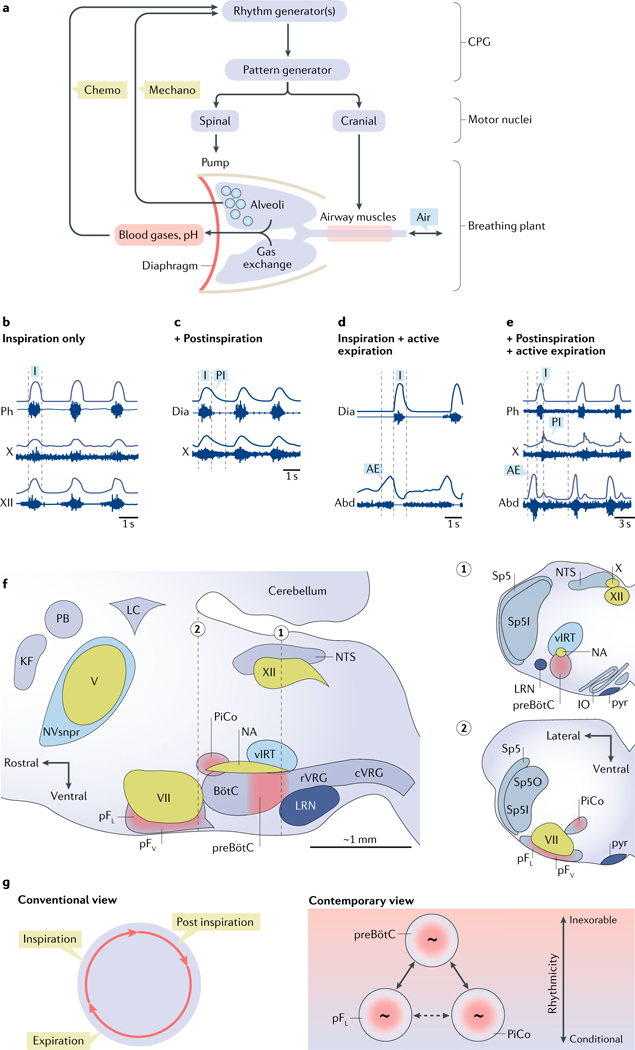
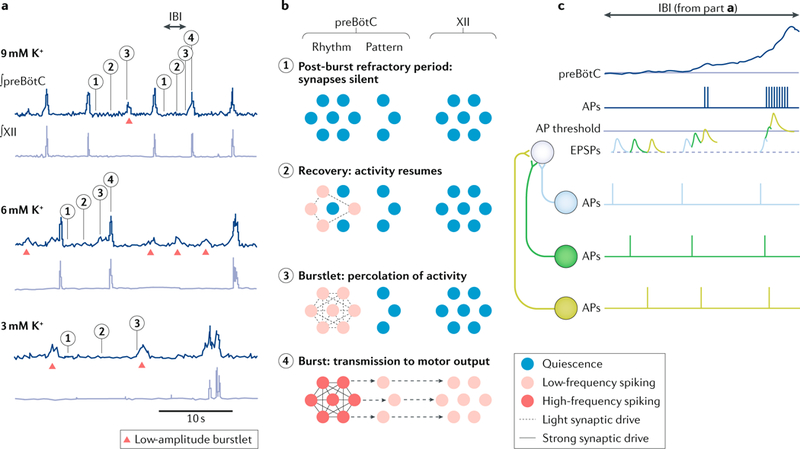
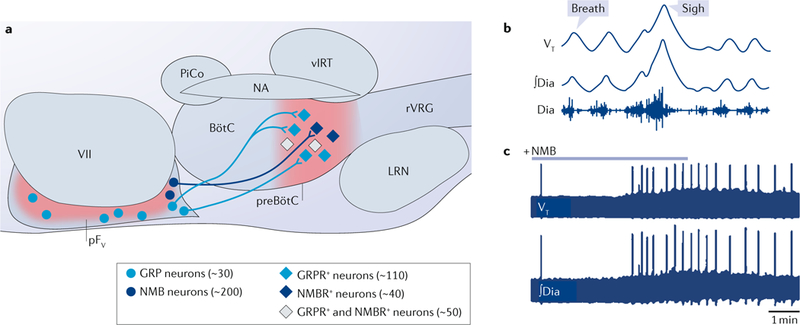
Breathing is a well-described, vital and surprisingly complex behaviour, with behavioural and physiological outputs that are easy to directly measure. Key neural elements for generating breathing pattern are distinct, compact and form a network amenable to detailed interrogation, promising the imminent discovery of molecular, cellular, synaptic and network mechanisms that give rise to the behaviour. Coupled oscillatory microcircuits make up the rhythmic core of the breathing network. Primary among these is the preBötzinger Complex (preBötC), which is composed of excitatory rhythmogenic interneurons and excitatory and inhibitory pattern-forming interneurons that together produce the essential periodic drive for inspiration. The preBötC coordinates all phases of the breathing cycle, coordinates breathing with orofacial behaviours and strongly influences, and is influenced by, emotion and cognition. Here, we review progress towards cracking the inner workings of this vital core.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
-
- Schwarzacher SW, Smith JC & Richter DW Pre-Bötzinger complex in the cat. J. Neurophysiol. 73, 1452–1461 (1995). - PubMed
-
- Schwarzacher SW, Rüb U & Deller T Neuroanatomical characteristics of the human pre-Bötzinger Complex and its involvement in neurodegenerative brainstem diseases. Brain J. Neurol. 134, 24–35 (2011). - PubMed
-
- Wenninger JM et al. Large lesions in the pre- Bötzinger complex area eliminate eupneic respiratory rhythm in awake goats. J. Appl. Physiol. 97, 1629–1636 (2004). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
