

Distribution of Initiation Times Reveals Mechanisms of Transcriptional Regulation in Single Cells
- PMID: 29742401
- PMCID: PMC5961459
- DOI: 10.1016/j.bpj.2018.03.031
Distribution of Initiation Times Reveals Mechanisms of Transcriptional Regulation in Single Cells
Abstract
Transcription is the dominant point of control of gene expression. Biochemical studies have revealed key molecular components of transcription and their interactions, but the dynamics of transcription initiation in cells is still poorly understood. This state of affairs is being remedied with experiments that observe transcriptional dynamics in single cells using fluorescent reporters. Quantitative information about transcription initiation dynamics can also be extracted from experiments that use electron micrographs of RNA polymerases caught in the act of transcribing a gene (Miller spreads). Inspired by these data, we analyze a general stochastic model of transcription initiation and elongation and compute the distribution of transcription initiation times. We show that different mechanisms of initiation leave distinct signatures in the distribution of initiation times that can be compared to experiments. We analyze published data from micrographs of RNA polymerases transcribing ribosomal RNA genes in Escherichia coli and compare the observed distributions of interpolymerase distances with the predictions from previously hypothesized mechanisms for the regulation of these genes. Our analysis demonstrates the potential of measuring the distribution of time intervals between initiation events as a probe for dissecting mechanisms of transcription initiation in live cells.
Copyright © 2018 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
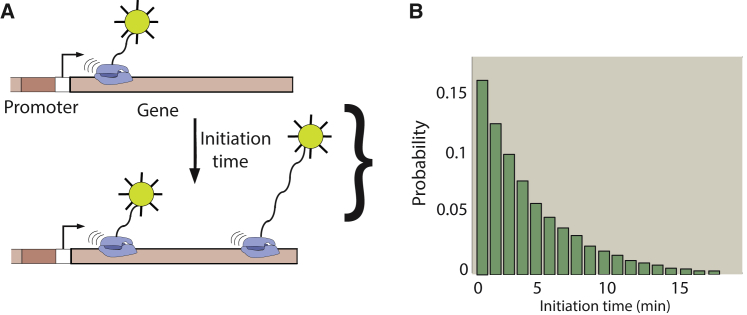
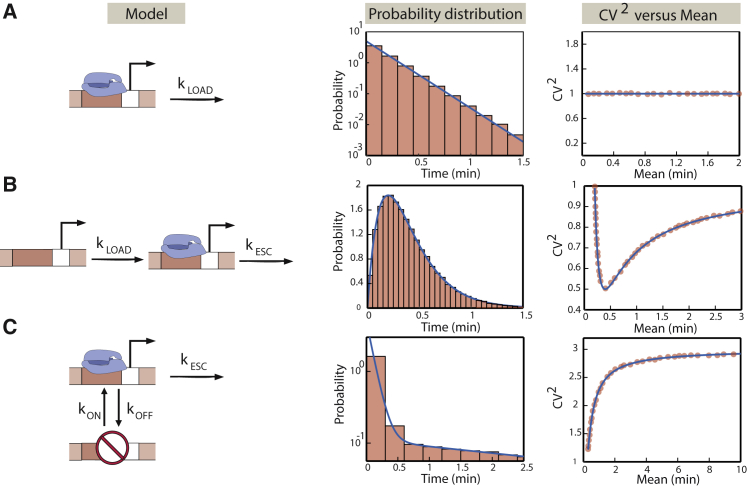
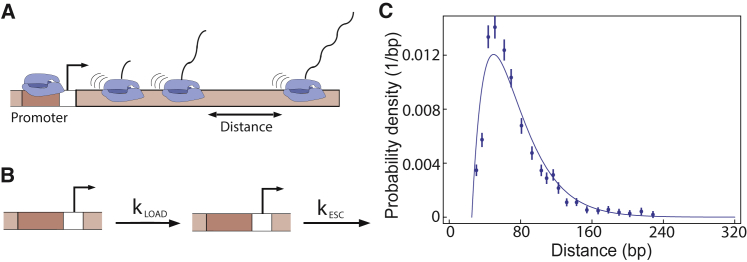
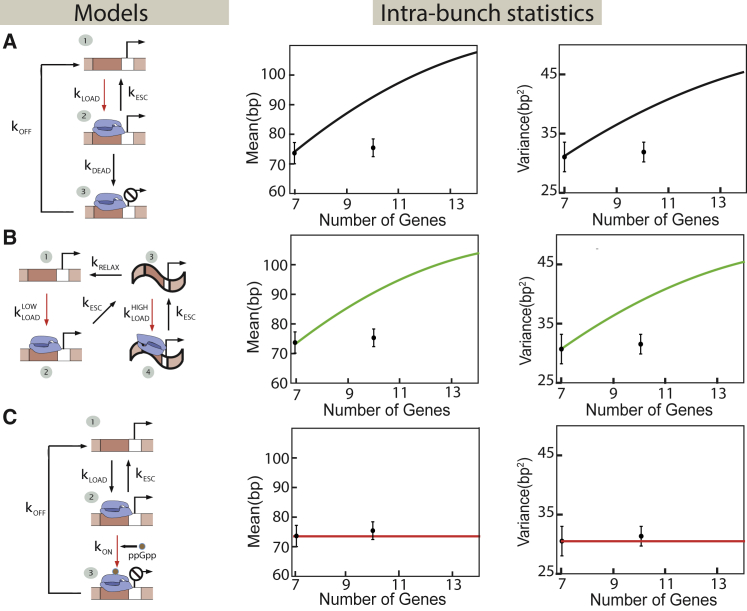
Similar articles
-
Probing Mechanisms of Transcription Elongation Through Cell-to-Cell Variability of RNA Polymerase.Biophys J. 2020 Apr 7;118(7):1769-1781. doi: 10.1016/j.bpj.2020.02.002. Epub 2020 Feb 12. Biophys J. 2020. PMID: 32101716 Free PMC article.
-
Increased rrn gene dosage causes intermittent transcription of rRNA in Escherichia coli.J Bacteriol. 1999 Jul;181(14):4170-5. doi: 10.1128/JB.181.14.4170-4175.1999. J Bacteriol. 1999. PMID: 10400572 Free PMC article.
-
The bacterial DNA-binding protein H-NS represses ribosomal RNA transcription by trapping RNA polymerase in the initiation complex.J Mol Biol. 2000 May 19;298(5):737-48. doi: 10.1006/jmbi.2000.3708. J Mol Biol. 2000. PMID: 10801345
-
Escherichia coli ribosomal RNA transcription: regulatory roles for ppGpp, NTPs, architectural proteins and a polymerase-binding protein.Mol Microbiol. 2005 Feb;55(4):973-7. doi: 10.1111/j.1365-2958.2004.04455.x. Mol Microbiol. 2005. PMID: 15686546 Review.
-
Transcription factor dynamics.Microbiology (Reading). 2008 Jul;154(Pt 7):1837-1844. doi: 10.1099/mic.0.2008/018549-0. Microbiology (Reading). 2008. PMID: 18599813 Review.
Cited by
-
RNA polymerase sliding on DNA can couple the transcription of nearby bacterial operons.bioRxiv [Preprint]. 2023 Feb 10:2023.02.10.528045. doi: 10.1101/2023.02.10.528045. bioRxiv. 2023. Update in: Proc Natl Acad Sci U S A. 2023 Jul 25;120(30):e2301402120. doi: 10.1073/pnas.2301402120. PMID: 36798213 Free PMC article. Updated. Preprint.
-
RNA polymerase sliding on DNA can couple the transcription of nearby bacterial operons.Proc Natl Acad Sci U S A. 2023 Jul 25;120(30):e2301402120. doi: 10.1073/pnas.2301402120. Epub 2023 Jul 17. Proc Natl Acad Sci U S A. 2023. PMID: 37459525 Free PMC article.
-
Constraining the complexity of promoter dynamics using fluctuations in gene expression.Phys Biol. 2019 Nov 5;17(1):015001. doi: 10.1088/1478-3975/ab4e57. Phys Biol. 2019. PMID: 31618721 Free PMC article.
-
Real-time single-cell characterization of the eukaryotic transcription cycle reveals correlations between RNA initiation, elongation, and cleavage.PLoS Comput Biol. 2021 May 18;17(5):e1008999. doi: 10.1371/journal.pcbi.1008999. eCollection 2021 May. PLoS Comput Biol. 2021. PMID: 34003867 Free PMC article.
-
Kinetic sculpting of the seven stripes of the Drosophila even-skipped gene.Elife. 2020 Dec 10;9:e61635. doi: 10.7554/eLife.61635. Elife. 2020. PMID: 33300492 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
