

Histidine168 is crucial for ΔpH-dependent gating of the human voltage-gated proton channel, hHV1
- PMID: 29743300
- PMCID: PMC5987877
- DOI: 10.1085/jgp.201711968
Histidine168 is crucial for ΔpH-dependent gating of the human voltage-gated proton channel, hHV1
Abstract
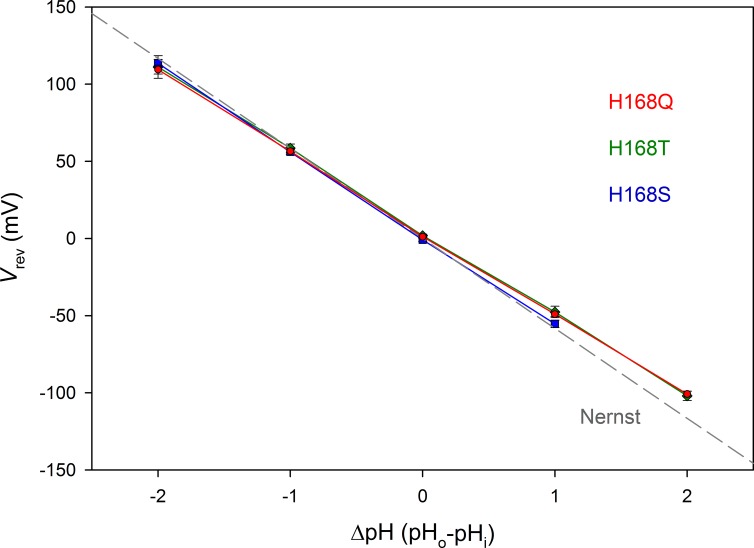
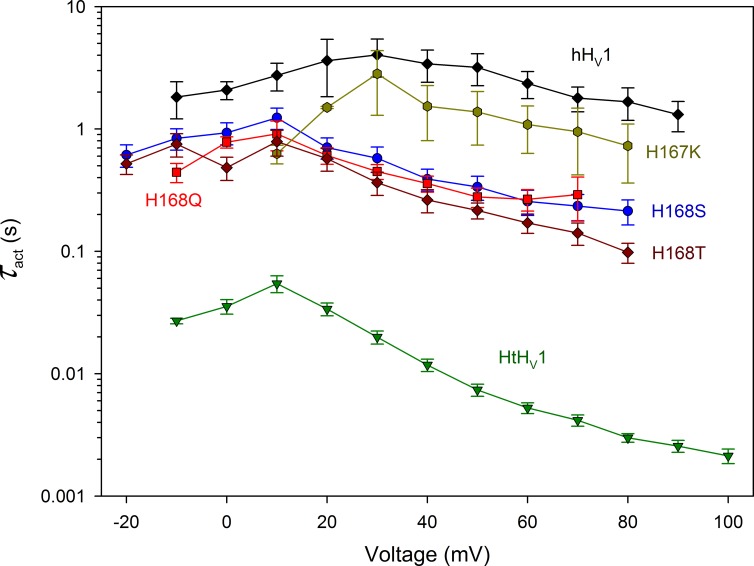
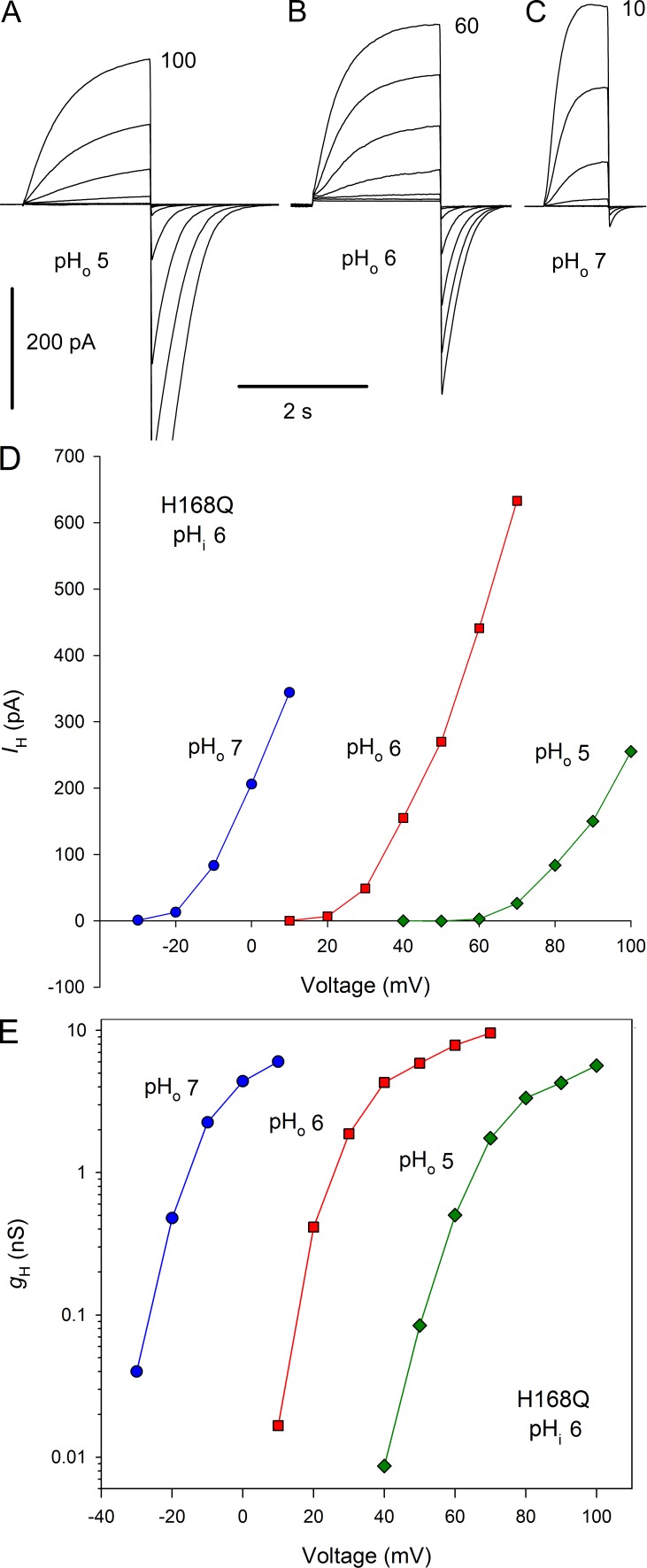
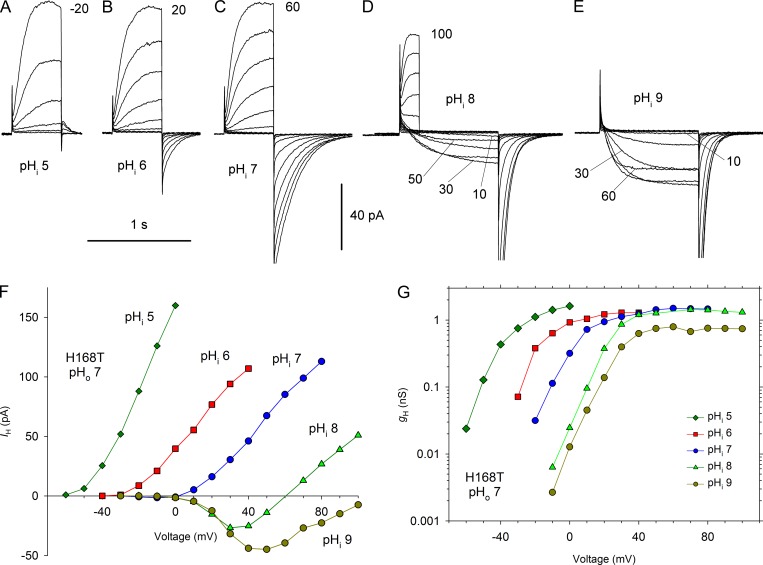
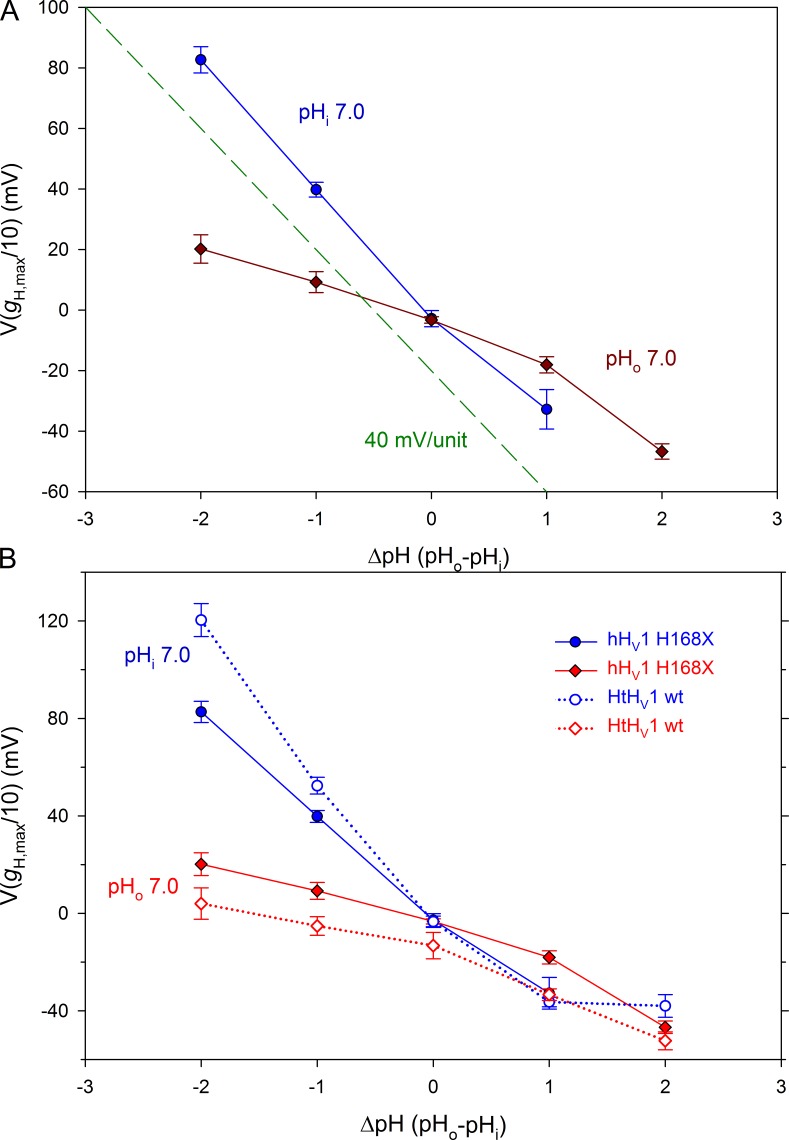
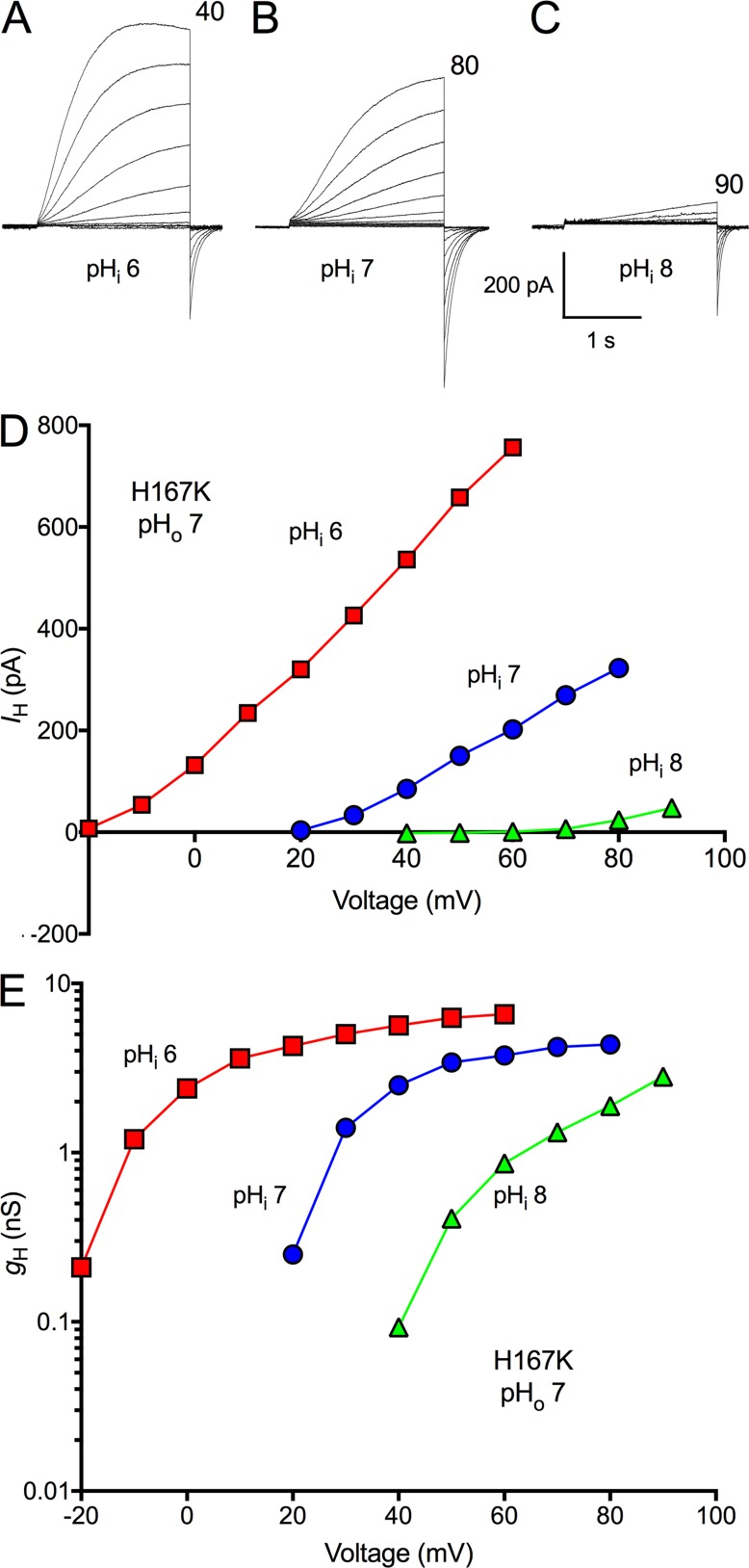
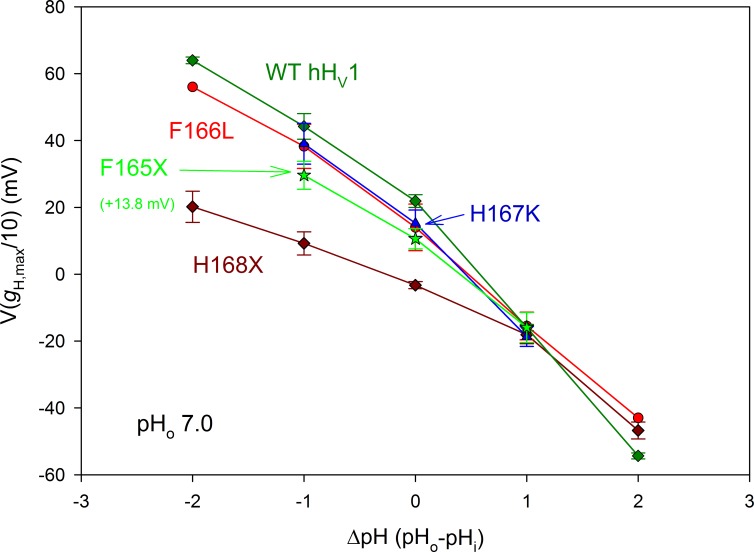
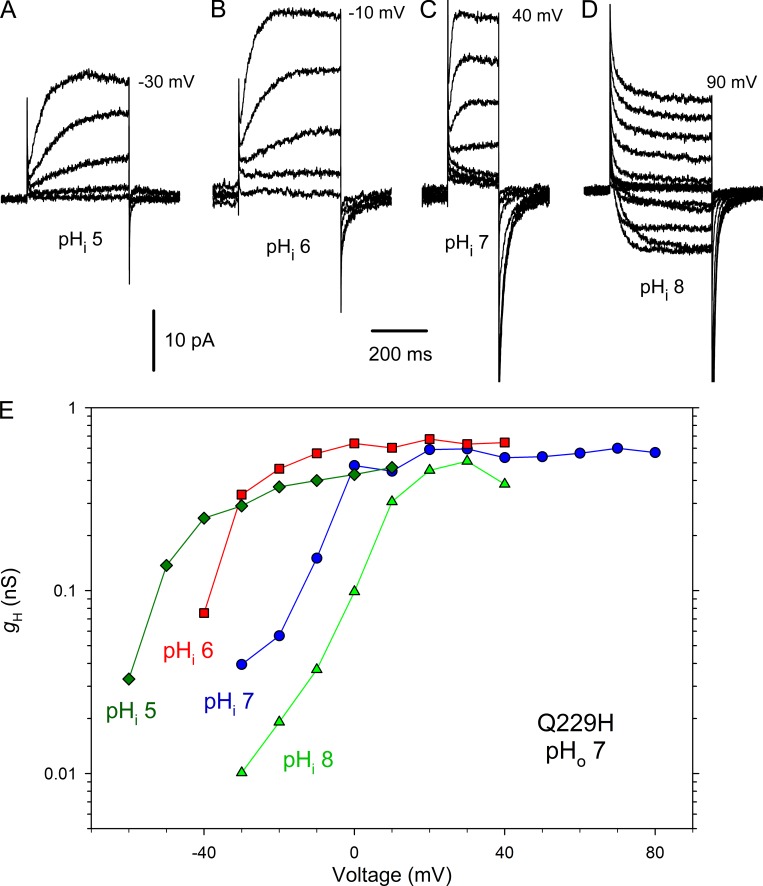
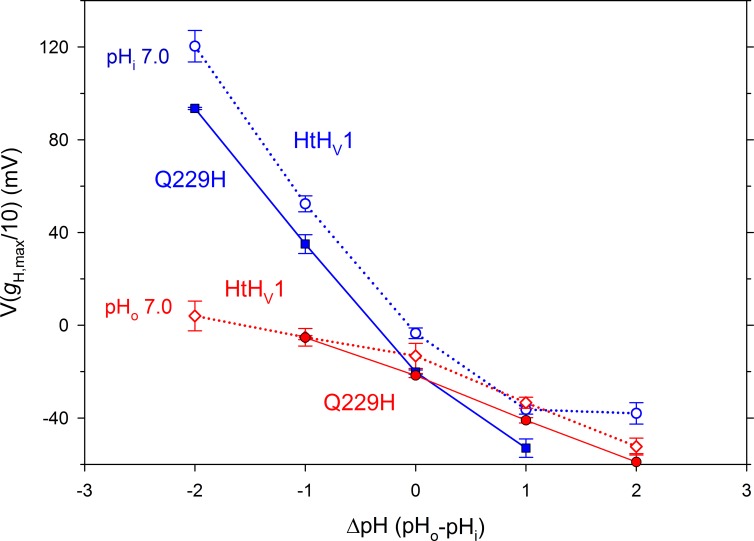
We recently identified a voltage-gated proton channel gene in the snail Helisoma trivolvis, HtHV1, and determined its electrophysiological properties. Consistent with early studies of proton currents in snail neurons, HtHV1 opens rapidly, but it unexpectedly exhibits uniquely defective sensitivity to intracellular pH (pHi). The H+ conductance (gH)-V relationship in the voltage-gated proton channel (HV1) from other species shifts 40 mV when either pHi or pHo (extracellular pH) is changed by 1 unit. This property, called ΔpH-dependent gating, is crucial to the functions of HV1 in many species and in numerous human tissues. The HtHV1 channel exhibits normal pHo dependence but anomalously weak pHi dependence. In this study, we show that a single point mutation in human hHV1-changing His168 to Gln168, the corresponding residue in HtHV1-compromises the pHi dependence of gating in the human channel so that it recapitulates the HtHV1 response. This location was previously identified as a contributor to the rapid gating kinetics of HV1 in Strongylocentrotus purpuratus His168 mutation in human HV1 accelerates activation but accounts for only a fraction of the species difference. H168Q, H168S, or H168T mutants exhibit normal pHo dependence, but changing pHi shifts the gH-V relationship on average by <20 mV/unit. Thus, His168 is critical to pHi sensing in hHV1. His168, located at the inner end of the pore on the S3 transmembrane helix, is the first residue identified in HV1 that significantly impairs pH sensing when mutated. Because pHo dependence remains intact, the selective erosion of pHi dependence supports the idea that there are distinct internal and external pH sensors. Although His168 may itself be a pHi sensor, the converse mutation, Q229H, does not normalize the pHi sensitivity of the HtHV1 channel. We hypothesize that the imidazole group of His168 interacts with nearby Phe165 or other parts of hHV1 to transduce pHi into shifts of voltage-dependent gating.
© 2018 Cherny et al.
Figures

Comment in
-
The acid test for pH-dependent gating in cloned HV1 channels.J Gen Physiol. 2018 Jun 4;150(6):781-782. doi: 10.1085/jgp.201812076. Epub 2018 May 15. J Gen Physiol. 2018. PMID: 29764925 Free PMC article.
References
-
- Berger T.K., Fußhöller D.M., Goodwin N., Bönigk W., Müller A., Dokani Khesroshahi N., Brenker C., Wachten D., Krause E., Kaupp U.B., and Strünker T.. 2017. Post-translational cleavage of Hv1 in human sperm tunes pH- and voltage-dependent gating. J. Physiol. 595:1533–1546. 10.1113/JP273189 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
