

Novel Strategy To Adapt Simian-Human Immunodeficiency Virus E1 Carrying env from an RV144 Volunteer to Rhesus Macaques: Coreceptor Switch and Final Recovery of a Pathogenic Virus with Exclusive R5 Tropism
- PMID: 29743361
- PMCID: PMC6026739
- DOI: 10.1128/JVI.02222-17
Novel Strategy To Adapt Simian-Human Immunodeficiency Virus E1 Carrying env from an RV144 Volunteer to Rhesus Macaques: Coreceptor Switch and Final Recovery of a Pathogenic Virus with Exclusive R5 Tropism
Abstract
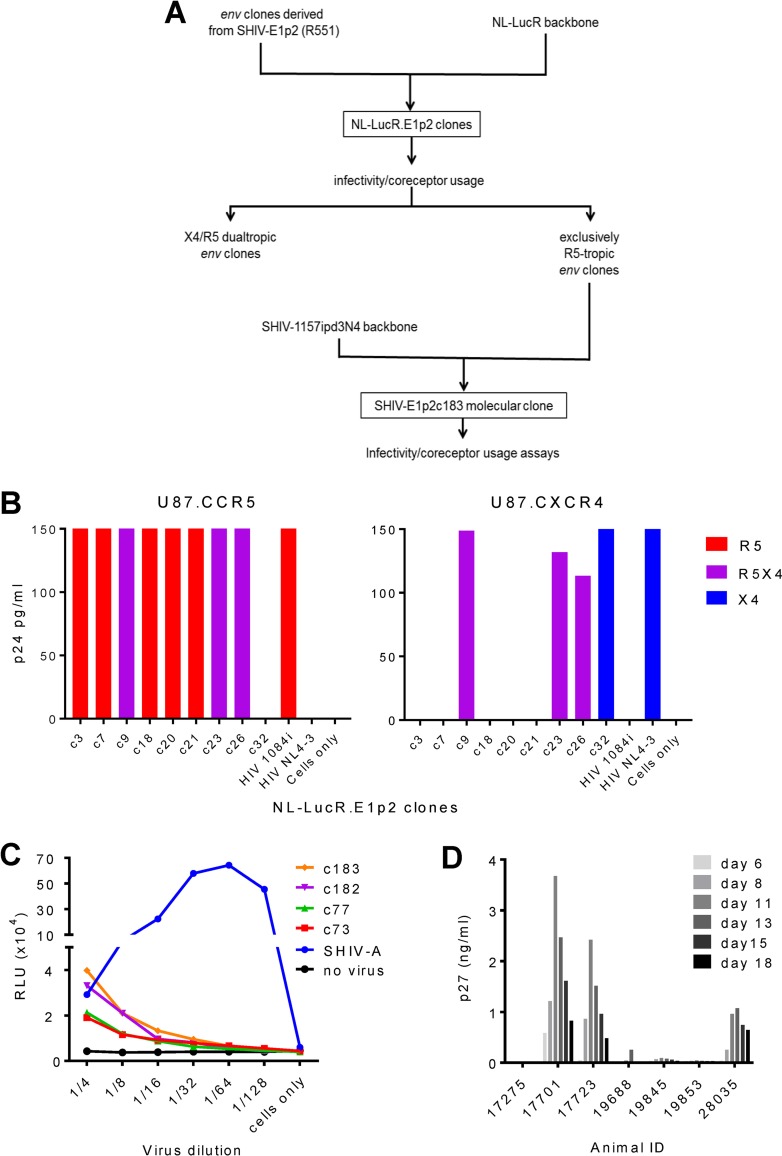
The phase III RV144 human immunodeficiency virus (HIV) vaccine trial conducted in Thailand remains the only study to show efficacy in decreasing the HIV acquisition risk. In Thailand, circulating recombinant forms of HIV clade A/E (CRF01_AE) predominate; in such viruses, env originates from clade E (HIV-E). We constructed a simian-human immunodeficiency virus (SHIV) chimera carrying env isolated from an RV144 placebo recipient in the SHIV-1157ipd3N4 backbone. The latter contains long terminal repeats (LTRs) with duplicated NF-κB sites, thus resembling HIV LTRs. We devised a novel strategy to adapt the parental infectious molecular clone (IMC), R5 SHIV-E1, to rhesus macaques: the simultaneous depletion of B and CD8+ cells followed by the intramuscular inoculation of proviral DNA and repeated administrations of cell-free virus. High-level viremia and CD4+ T-cell depletion ensued. Passage 3 virus unexpectedly caused acute, irreversible CD4+ T-cell loss; the partially adapted SHIV had become dual tropic. Virus and IMCs with exclusive R5 tropism were reisolated from earlier passages, combined, and used to complete adaptation through additional macaques. The final isolate, SHIV-E1p5, remained solely R5 tropic. It had a tier 2 neutralization phenotype, was mucosally transmissible, and was pathogenic. Deep sequencing revealed 99% Env amino acid sequence conservation; X4-only and dual-tropic strains had evolved independently from an early branch of parental SHIV-E1. To conclude, our primate model data reveal that SHIV-E1p5 recapitulates important aspects of HIV transmission and pathobiology in humans.IMPORTANCE Understanding the protective principles that lead to a safe, effective vaccine against HIV in nonhuman primate (NHP) models requires test viruses that allow the evaluation of anti-HIV envelope responses. Reduced HIV acquisition risk in RV144 has been linked to nonneutralizing IgG antibodies with a range of effector activities. Definitive experiments to decipher the mechanisms of the partial protection observed in RV144 require passive-immunization studies in NHPs with a relevant test virus. We have generated such a virus by inserting env from an RV144 placebo recipient into a SHIV backbone with HIV-like LTRs. The final SHIV-E1p5 isolate, grown in rhesus monkey peripheral blood mononuclear cells, was mucosally transmissible and pathogenic. Earlier SHIV-E passages showed a coreceptor switch, again mimicking HIV biology in humans. Thus, our series of SHIV-E strains mirrors HIV transmission and disease progression in humans. SHIV-E1p5 represents a biologically relevant tool to assess prevention strategies.
Keywords: CRF01_AE; HIV; RV144 trial; SHIV-E; adaptation; coreceptor switch; rhesus macaques.
Copyright © 2018 American Society for Microbiology.
Figures
Similar articles
-
Molecularly cloned SHIV-1157ipd3N4: a highly replication- competent, mucosally transmissible R5 simian-human immunodeficiency virus encoding HIV clade C Env.J Virol. 2006 Sep;80(17):8729-38. doi: 10.1128/JVI.00558-06. J Virol. 2006. PMID: 16912320 Free PMC article.
-
Simian-human immunodeficiency virus containing a human immunodeficiency virus type 1 subtype-E envelope gene: persistent infection, CD4(+) T-cell depletion, and mucosal membrane transmission in macaques.J Virol. 2000 Sep;74(17):7851-60. doi: 10.1128/jvi.74.17.7851-7860.2000. J Virol. 2000. PMID: 10933692 Free PMC article.
-
Enhanced infectivity of an R5-tropic simian/human immunodeficiency virus carrying human immunodeficiency virus type 1 subtype C envelope after serial passages in pig-tailed macaques (Macaca nemestrina).J Virol. 2000 Jul;74(14):6501-10. doi: 10.1128/jvi.74.14.6501-6510.2000. J Virol. 2000. PMID: 10864663 Free PMC article.
-
Understanding the basis of CD4(+) T-cell depletion in macaques infected by a simian-human immunodeficiency virus.Vaccine. 2002 May 6;20(15):1934-7. doi: 10.1016/s0264-410x(02)00072-5. Vaccine. 2002. PMID: 11983249 Review.
-
The SIV Envelope Glycoprotein, Viral Tropism, and Pathogenesis: Novel Insights from Nonhuman Primate Models of AIDS.Curr HIV Res. 2018;16(1):29-40. doi: 10.2174/1570162X15666171124123116. Curr HIV Res. 2018. PMID: 29173176 Review.
Cited by
-
Adaptation of an R5 Simian-Human Immunodeficiency Virus Encoding an HIV Clade A Envelope with or without Ablation of Adaptive Host Immunity: Differential Selection of Viral Mutants.J Virol. 2019 Apr 17;93(9):e02267-18. doi: 10.1128/JVI.02267-18. Print 2019 May 1. J Virol. 2019. PMID: 30760566 Free PMC article.
-
Host immunity associated with spontaneous suppression of viremia in therapy-naïve young rhesus macaques following neonatal SHIV infection.J Virol. 2023 Nov 30;97(11):e0109423. doi: 10.1128/jvi.01094-23. Epub 2023 Oct 24. J Virol. 2023. PMID: 37874153 Free PMC article.
References
-
- Rerks-Ngarm S, Pitisuttithum P, Nitayaphan S, Kaewkungwal J, Chiu J, Paris R, Premsri N, Namwat C, de Souza M, Adams E, Benenson M, Gurunathan S, Tartaglia J, McNeil JG, Francis DP, Stablein D, Birx DL, Chunsuttiwat S, Khamboonruang C, Thongcharoen P, Robb ML, Michael NL, Kunasol P, Kim JH. 2009. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N Engl J Med 361:2209–2220. doi: 10.1056/NEJMoa0908492. - DOI - PubMed
-
- Haynes BF, Gilbert PB, McElrath MJ, Zolla-Pazner S, Tomaras GD, Alam SM, Evans DT, Montefiori DC, Karnasuta C, Sutthent R, Liao HX, DeVico AL, Lewis GK, Williams C, Pinter A, Fong Y, Janes H, DeCamp A, Huang Y, Rao M, Billings E, Karasavvas N, Robb ML, Ngauy V, de Souza MS, Paris R, Ferrari G, Bailer RT, Soderberg KA, Andrews C, Berman PW, Frahm N, De Rosa SC, Alpert MD, Yates NL, Shen X, Koup RA, Pitisuttithum P, Kaewkungwal J, Nitayaphan S, Rerks-Ngarm S, Michael NL, Kim JH. 2012. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N Engl J Med 366:1275–1286. doi: 10.1056/NEJMoa1113425. - DOI - PMC - PubMed
-
- Bonsignori M, Pollara J, Moody MA, Alpert MD, Chen X, Hwang KK, Gilbert PB, Huang Y, Gurley TC, Kozink DM, Marshall DJ, Whitesides JF, Tsao CY, Kaewkungwal J, Nitayaphan S, Pitisuttithum P, Rerks-Ngarm S, Kim JH, Michael NL, Tomaras GD, Montefiori DC, Lewis GK, DeVico A, Evans DT, Ferrari G, Liao HX, Haynes BF. 2012. Antibody-dependent cellular cytotoxicity-mediating antibodies from an HIV-1 vaccine efficacy trial target multiple epitopes and preferentially use the VH1 gene family. J Virol 86:11521–11532. doi: 10.1128/JVI.01023-12. - DOI - PMC - PubMed
-
- Chung AW, Ghebremichael M, Robinson H, Brown E, Choi I, Lane S, Dugast AS, Schoen MK, Rolland M, Suscovich TJ, Mahan AE, Liao L, Streeck H, Andrews C, Rerks-Ngarm S, Nitayaphan S, de Souza MS, Kaewkungwal J, Pitisuttithum P, Francis D, Michael NL, Kim JH, Bailey-Kellogg C, Ackerman ME, Alter G. 2014. Polyfunctional Fc-effector profiles mediated by IgG subclass selection distinguish RV144 and VAX003 vaccines. Sci Transl Med 6:228ra38. doi: 10.1126/scitranslmed.3007736. - DOI - PubMed
-
- Chung AW, Kumar MP, Arnold KB, Yu WH, Schoen MK, Dunphy LJ, Suscovich TJ, Frahm N, Linde C, Mahan AE, Hoffner M, Streeck H, Ackerman ME, McElrath MJ, Schuitemaker H, Pau MG, Baden LR, Kim JH, Michael NL, Barouch DH, Lauffenburger DA, Alter G. 2015. Dissecting polyclonal vaccine-induced humoral immunity against HIV using systems serology. Cell 163:988–998. doi: 10.1016/j.cell.2015.10.027. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
