

Synaptic phospholipids as a new target for cortical hyperexcitability and E/I balance in psychiatric disorders
- PMID: 29743582
- PMCID: PMC6153268
- DOI: 10.1038/s41380-018-0053-1
Synaptic phospholipids as a new target for cortical hyperexcitability and E/I balance in psychiatric disorders
Erratum in
-
Correction: Synaptic phospholipids as a new target for cortical hyperexcitability and E/I balance in psychiatric disorders.Mol Psychiatry. 2020 Nov;25(11):3108. doi: 10.1038/s41380-018-0320-1. Mol Psychiatry. 2020. PMID: 30602735 Free PMC article.
Abstract
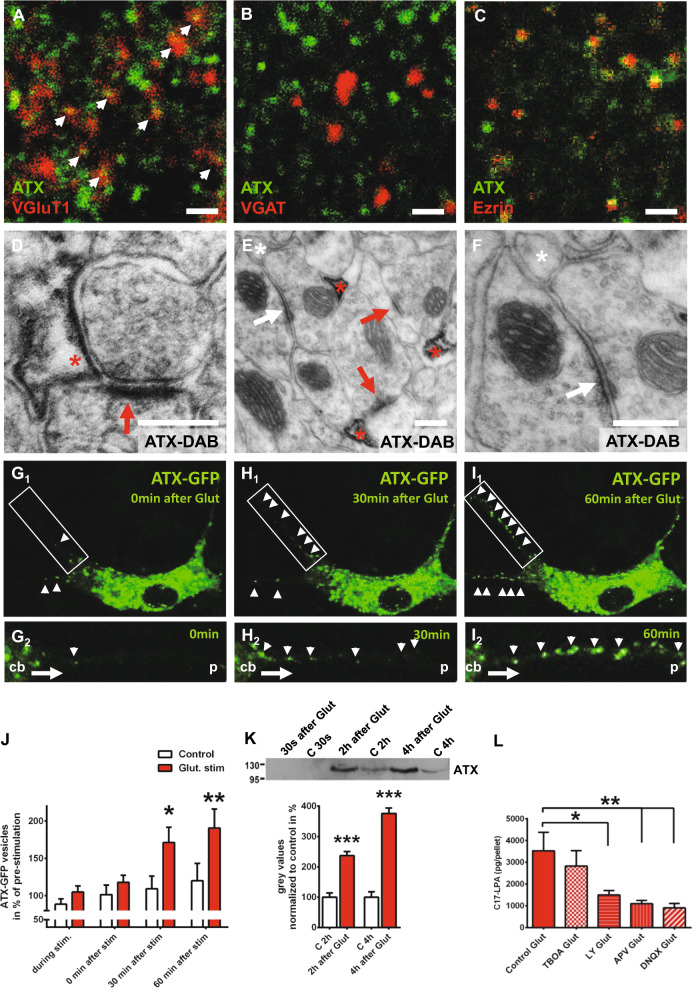
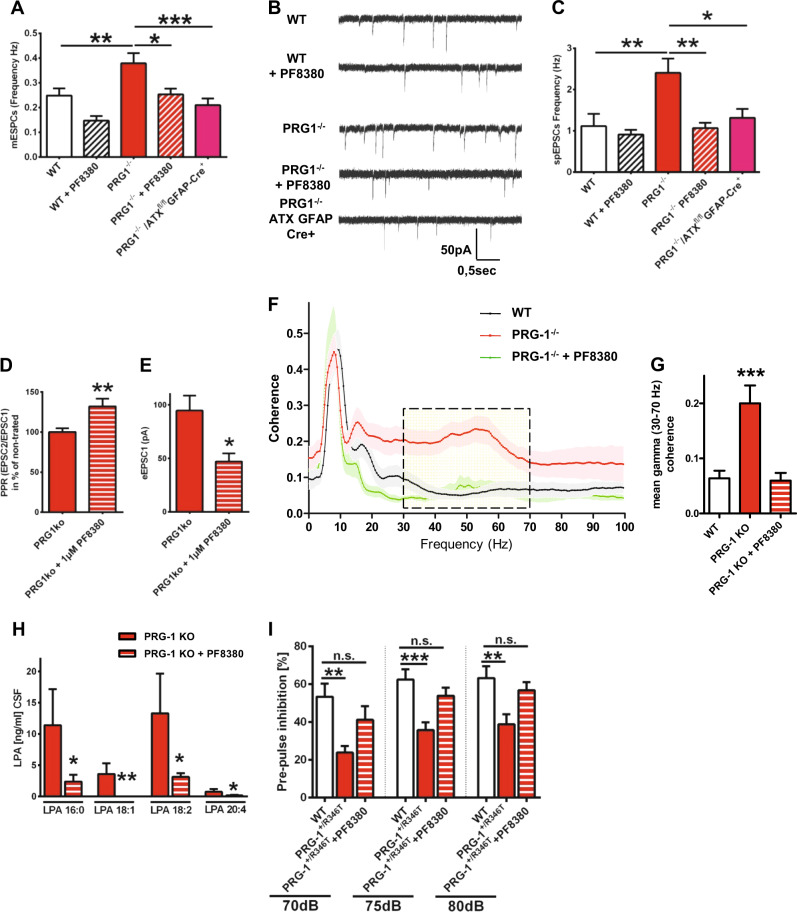
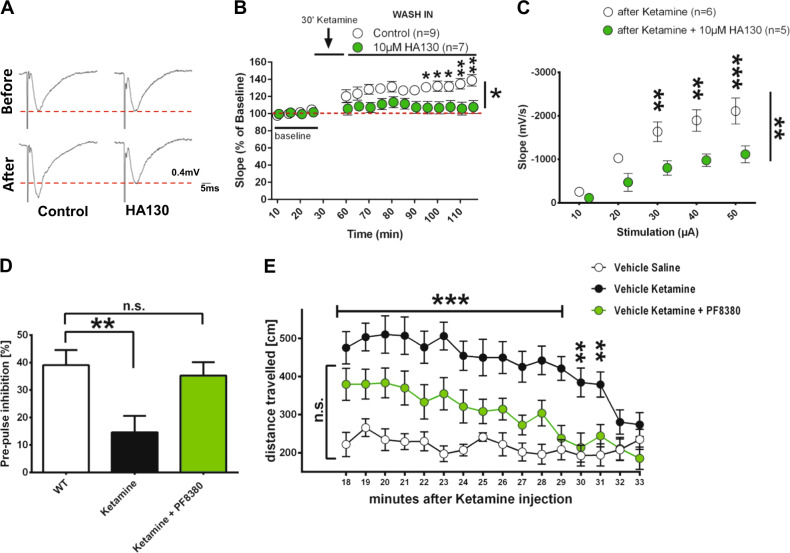
Lysophosphatidic acid (LPA) is a synaptic phospholipid, which regulates cortical excitation/inhibition (E/I) balance and controls sensory information processing in mice and man. Altered synaptic LPA signaling was shown to be associated with psychiatric disorders. Here, we show that the LPA-synthesizing enzyme autotaxin (ATX) is expressed in the astrocytic compartment of excitatory synapses and modulates glutamatergic transmission. In astrocytes, ATX is sorted toward fine astrocytic processes and transported to excitatory but not inhibitory synapses. This ATX sorting, as well as the enzymatic activity of astrocyte-derived ATX are dynamically regulated by neuronal activity via astrocytic glutamate receptors. Pharmacological and genetic ATX inhibition both rescued schizophrenia-related hyperexcitability syndromes caused by altered bioactive lipid signaling in two genetic mouse models for psychiatric disorders. Interestingly, ATX inhibition did not affect naive animals. However, as our data suggested that pharmacological ATX inhibition is a general method to reverse cortical excitability, we applied ATX inhibition in a ketamine model of schizophrenia and rescued thereby the electrophysiological and behavioral schizophrenia-like phenotype. Our data show that astrocytic ATX is a novel modulator of glutamatergic transmission and that targeting ATX might be a versatile strategy for a novel drug therapy to treat cortical hyperexcitability in psychiatric disorders.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
