

Identification of a transporter complex responsible for the cytosolic entry of nitrogen-containing bisphosphonates
- PMID: 29745899
- PMCID: PMC6021172
- DOI: 10.7554/eLife.36620
Identification of a transporter complex responsible for the cytosolic entry of nitrogen-containing bisphosphonates
Abstract
Nitrogen-containing-bisphosphonates (N-BPs) are a class of drugs widely prescribed to treat osteoporosis and other bone-related diseases. Although previous studies have established that N-BPs function by inhibiting the mevalonate pathway in osteoclasts, the mechanism by which N-BPs enter the cytosol from the extracellular space to reach their molecular target is not understood. Here, we implemented a CRISPRi-mediated genome-wide screen and identified SLC37A3 (solute carrier family 37 member A3) as a gene required for the action of N-BPs in mammalian cells. We observed that SLC37A3 forms a complex with ATRAID (all-trans retinoic acid-induced differentiation factor), a previously identified genetic target of N-BPs. SLC37A3 and ATRAID localize to lysosomes and are required for releasing N-BP molecules that have trafficked to lysosomes through fluid-phase endocytosis into the cytosol. Our results elucidate the route by which N-BPs are delivered to their molecular target, addressing a key aspect of the mechanism of action of N-BPs that may have significant clinical relevance.
Keywords: biochemistry; cell biology; chemical biology; genome-wide screening; human; lysosomes; mechanism of action; membrane transporter; mouse.
© 2018, Yu et al.
Conflict of interest statement
ZY, LS, CP, MH, GW, MA, TP, DS, JW No competing interests declared, EO Chief Scientific Officer and a Vice President at the Howard Hughes Medical Institute, one of the three founding funders of eLife.
Figures
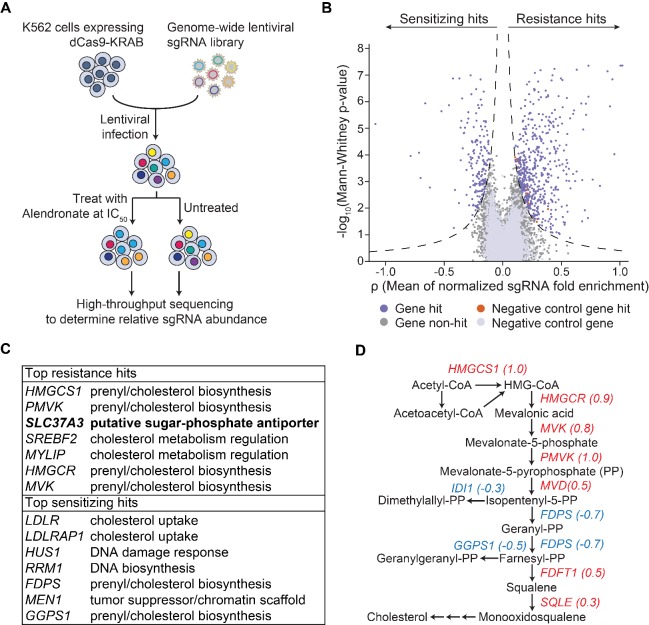
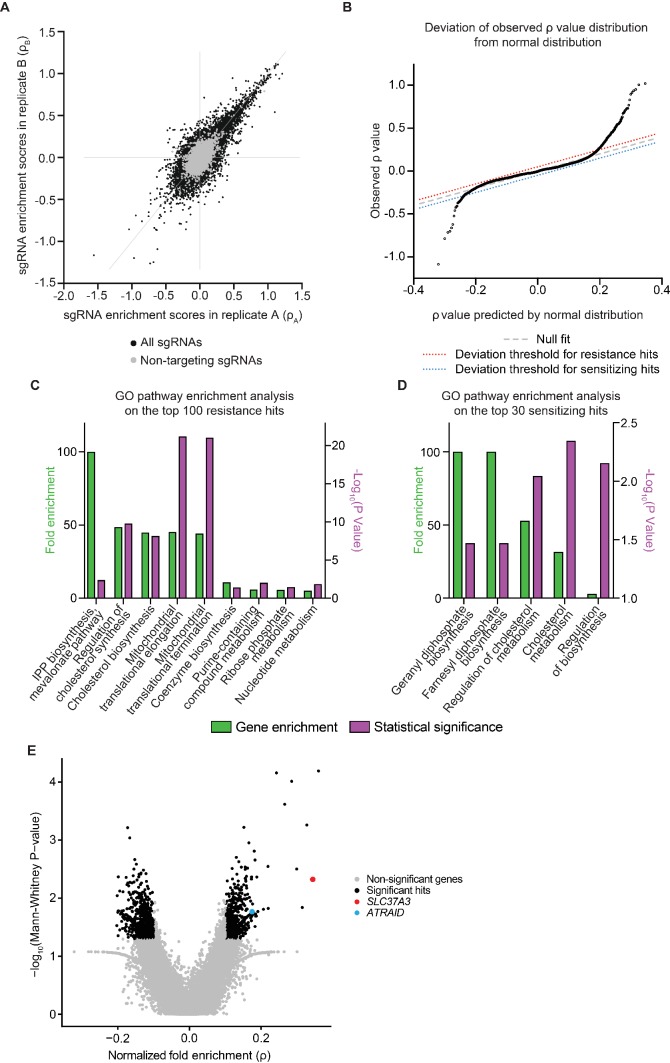
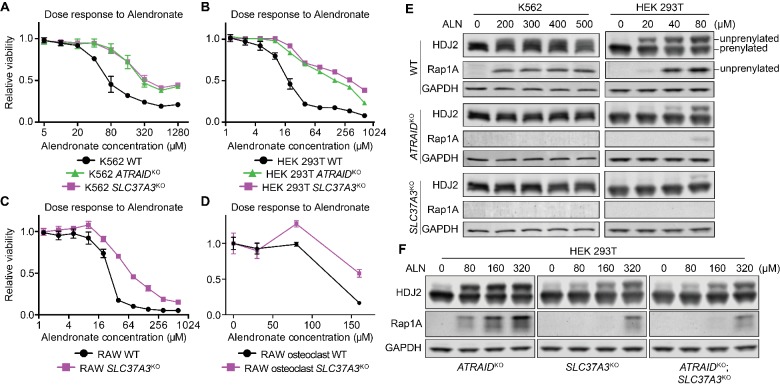
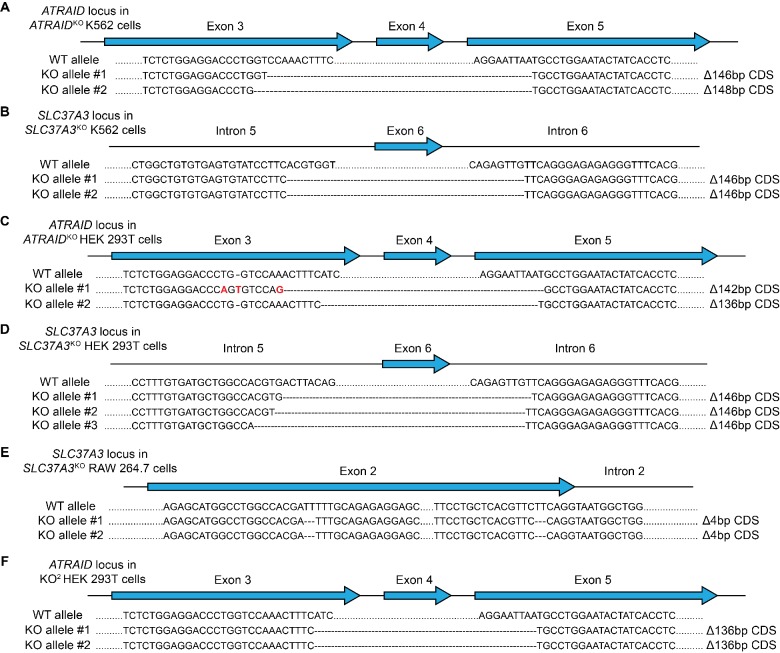

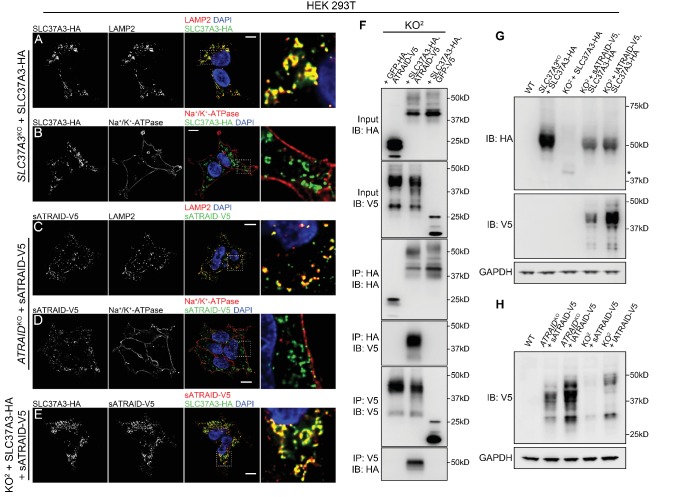

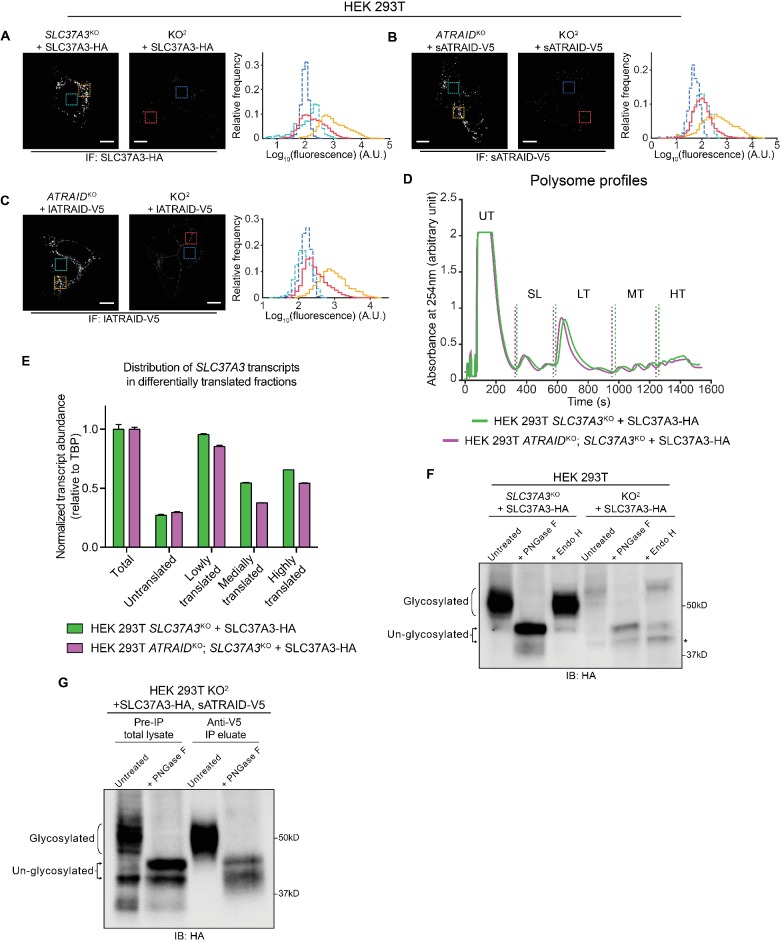
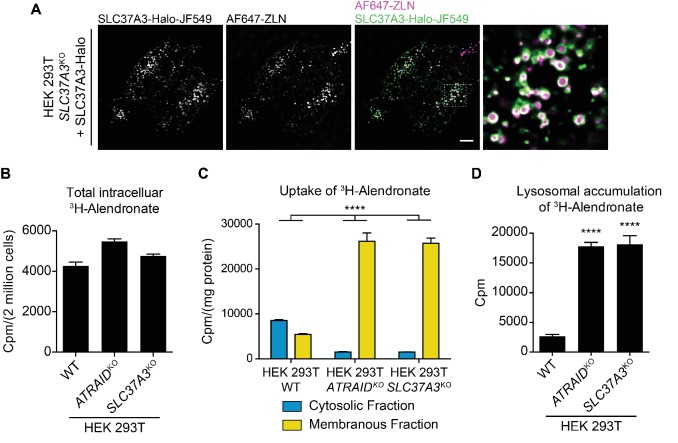
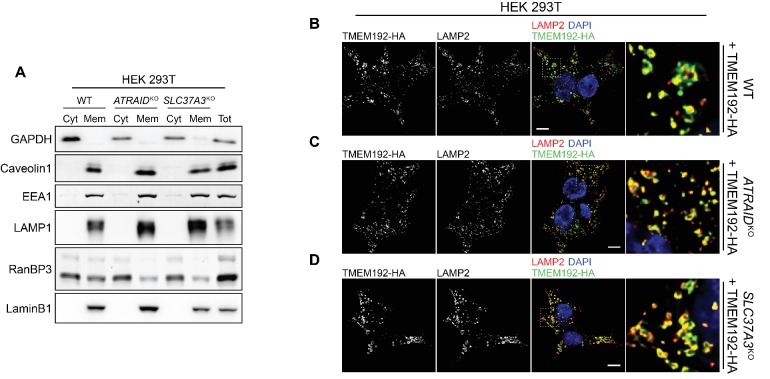
Comment in
-
From vesicle to cytosol.Elife. 2018 Jun 27;7:e38847. doi: 10.7554/eLife.38847. Elife. 2018. PMID: 29947329 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
