

A fluorescent reporter for quantification and enrichment of DNA editing by APOBEC-Cas9 or cleavage by Cas9 in living cells
- PMID: 29746667
- PMCID: PMC6101615
- DOI: 10.1093/nar/gky332
A fluorescent reporter for quantification and enrichment of DNA editing by APOBEC-Cas9 or cleavage by Cas9 in living cells
Abstract
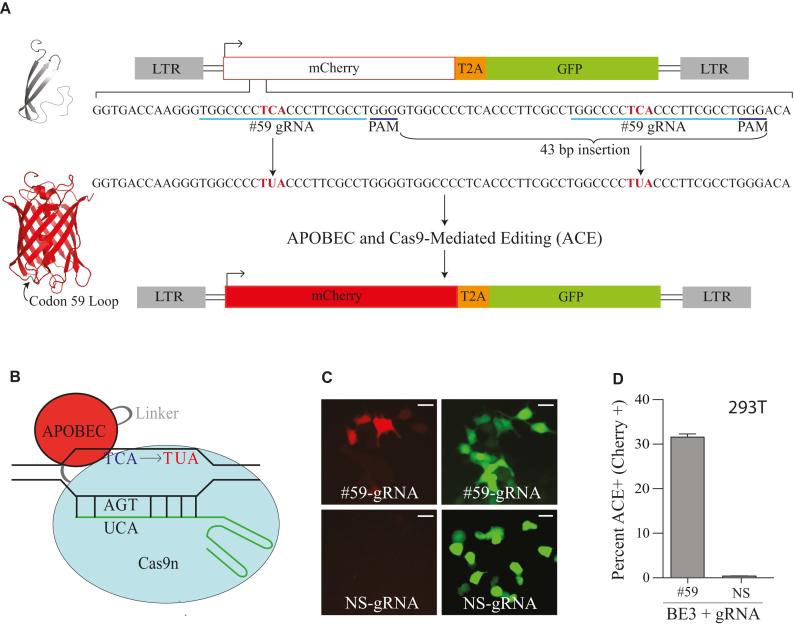
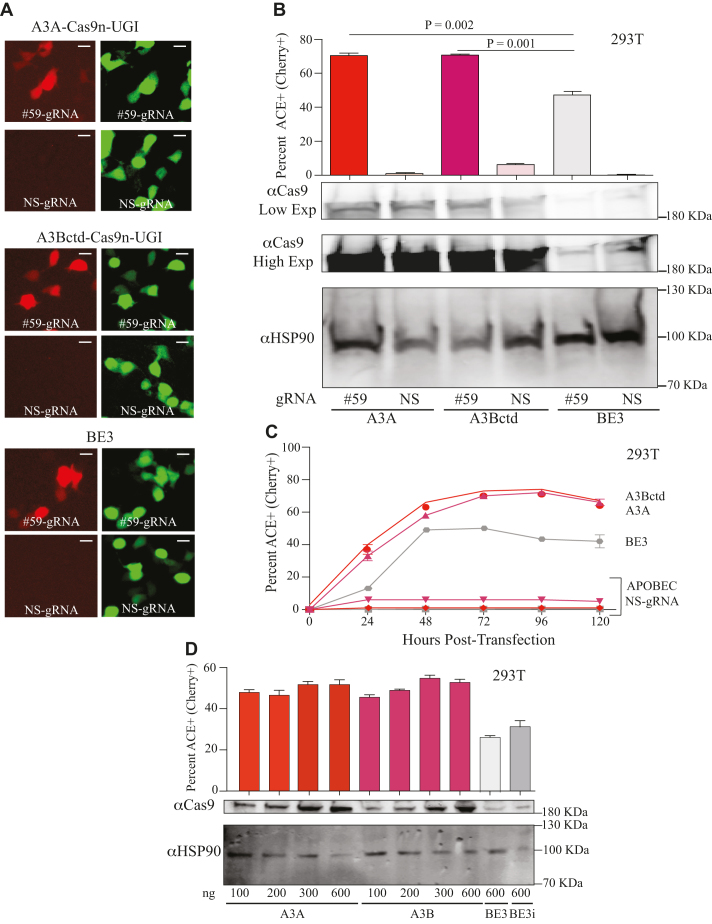
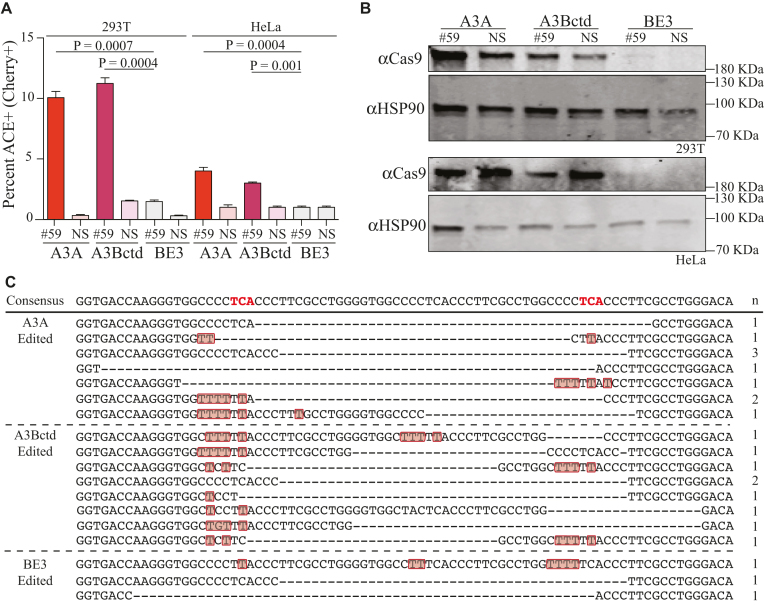
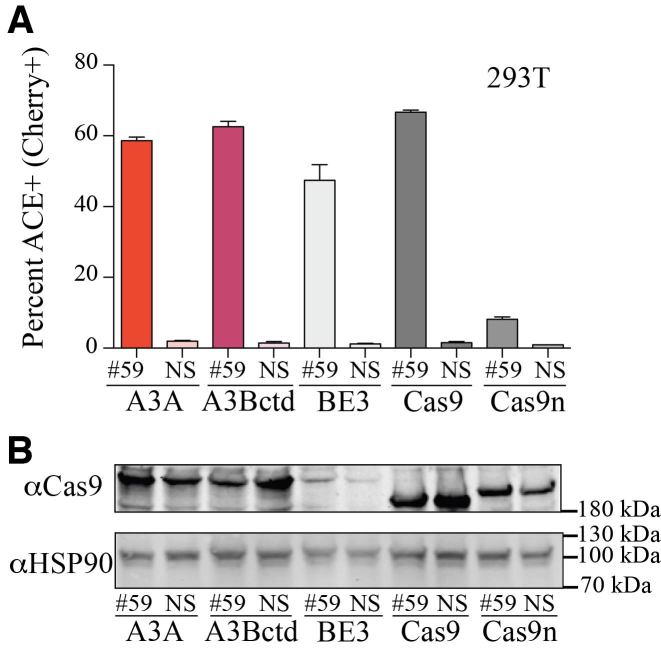
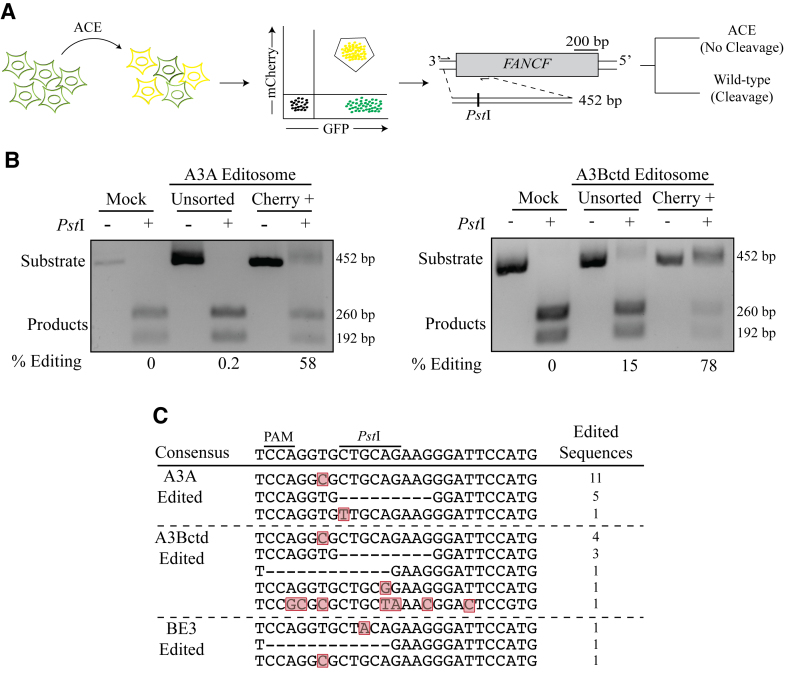
Base editing is an exciting new genome engineering technology. C-to-T mutations in genomic DNA have been achieved using ribonucleoprotein complexes comprised of rat APOBEC1 single-stranded DNA deaminase, Cas9 nickase (Cas9n), uracil DNA glycosylase inhibitor (UGI), and guide (g)RNA. Here, we report the first real-time reporter system for quantification of APOBEC-mediated base editing activity in living mammalian cells. The reporter expresses eGFP constitutively as a marker for transfection or transduction, and editing restores functionality of an upstream mCherry cassette through the simultaneous processing of two gRNA binding regions that each contain an APOBEC-preferred 5'TCA target site. Using this system as both an episomal and a chromosomal editing reporter, we show that human APOBEC3A-Cas9n-UGI and APOBEC3B-Cas9n-UGI base editing complexes are more efficient than the original rat APOBEC1-Cas9n-UGI construct. We also demonstrate coincident enrichment of editing events at a heterologous chromosomal locus in reporter-edited, mCherry-positive cells. The mCherry reporter also quantifies the double-stranded DNA cleavage activity of Cas9, and may therefore be adaptable for use with many different CRISPR systems. The combination of a rapid, fluorescence-based editing reporter system and more efficient, structurally defined DNA editing enzymes broadens the versatility of the rapidly expanding toolbox of genome editing and engineering technologies.
Figures
References
-
- Harris R.S., Petersen-Mahrt S.K., Neuberger M.S.. RNA editing enzyme APOBEC1 and some of its homologs can act as DNA mutators. Mol. Cell. 2002; 10:1247–1253. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
