

Spontaneous DNA damage to the nuclear genome promotes senescence, redox imbalance and aging
- PMID: 29747066
- PMCID: PMC6006678
- DOI: 10.1016/j.redox.2018.04.007
Spontaneous DNA damage to the nuclear genome promotes senescence, redox imbalance and aging
Abstract
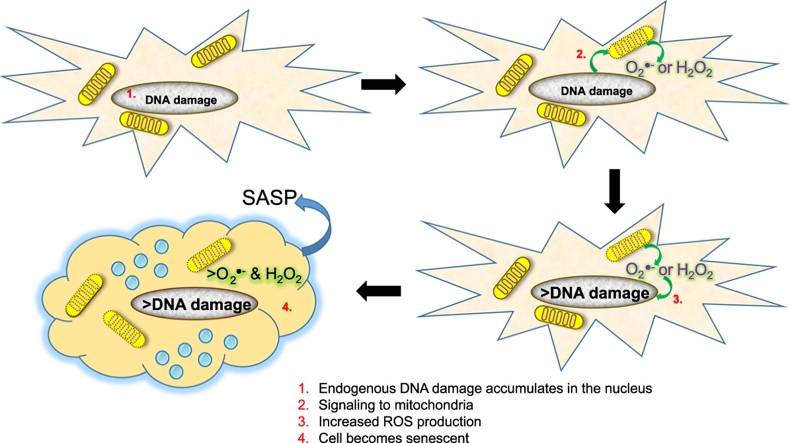
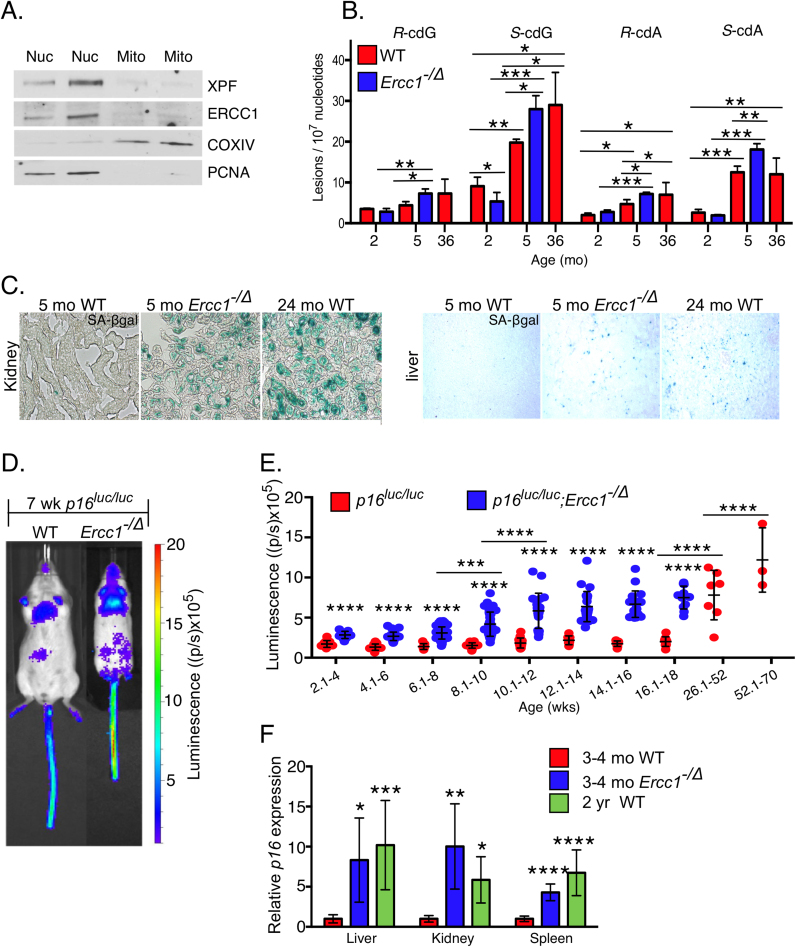
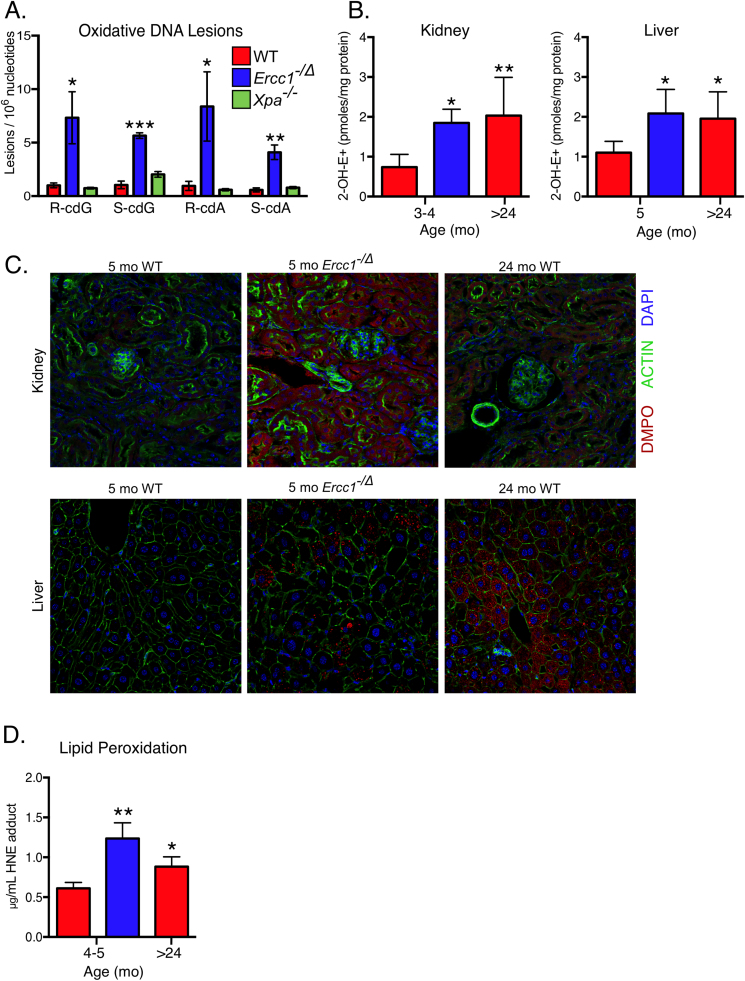
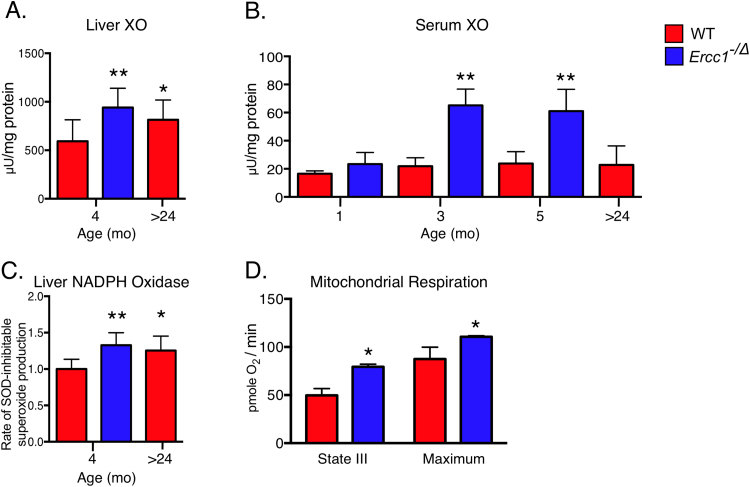
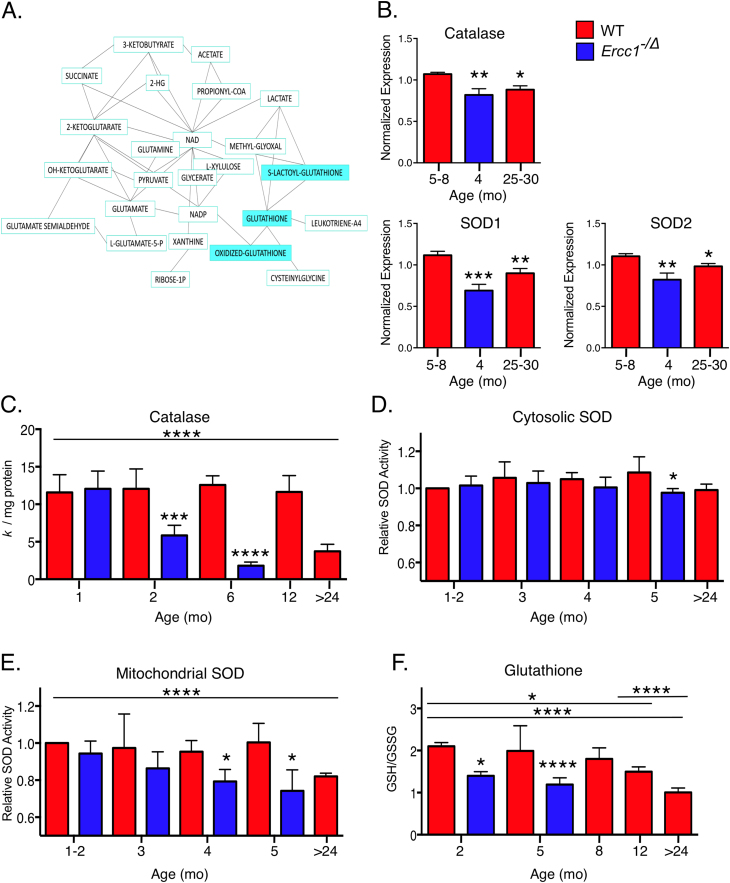
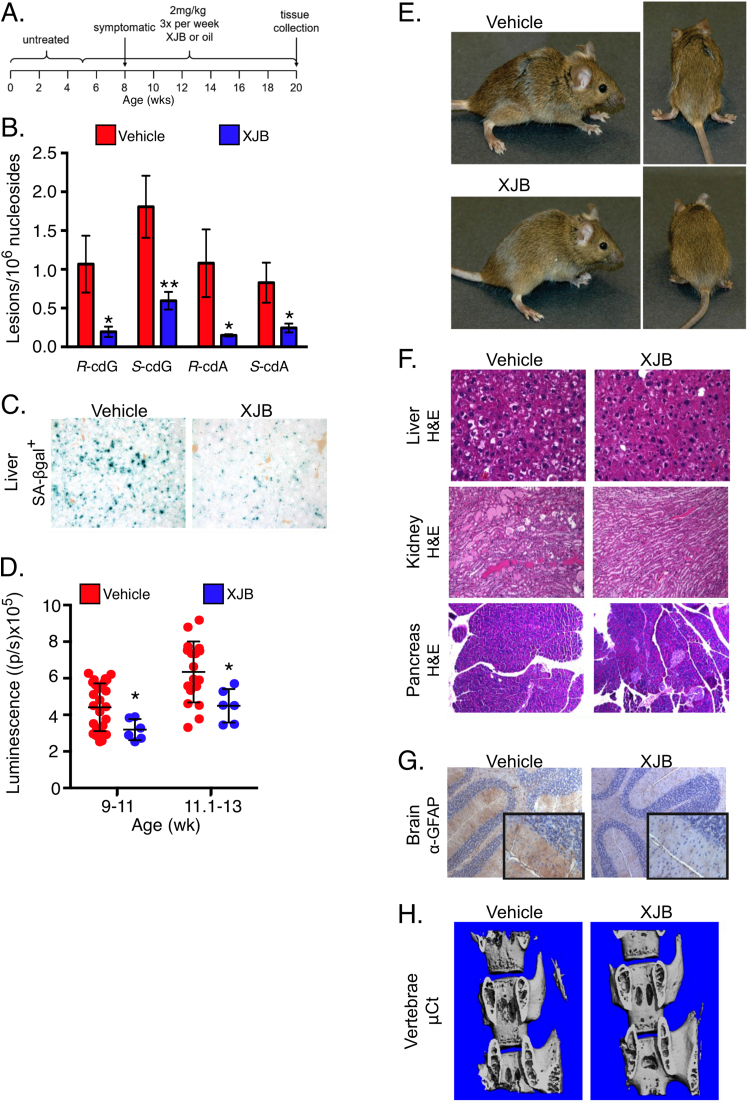
Accumulation of senescent cells over time contributes to aging and age-related diseases. However, what drives senescence in vivo is not clear. Here we used a genetic approach to determine if spontaneous nuclear DNA damage is sufficient to initiate senescence in mammals. Ercc1-/∆ mice with reduced expression of ERCC1-XPF endonuclease have impaired capacity to repair the nuclear genome. Ercc1-/∆ mice accumulated spontaneous, oxidative DNA damage more rapidly than wild-type (WT) mice. As a consequence, senescent cells accumulated more rapidly in Ercc1-/∆ mice compared to repair-competent animals. However, the levels of DNA damage and senescent cells in Ercc1-/∆ mice never exceeded that observed in old WT mice. Surprisingly, levels of reactive oxygen species (ROS) were increased in tissues of Ercc1-/∆ mice to an extent identical to naturally-aged WT mice. Increased enzymatic production of ROS and decreased antioxidants contributed to the elevation in oxidative stress in both Ercc1-/∆ and aged WT mice. Chronic treatment of Ercc1-/∆ mice with the mitochondrial-targeted radical scavenger XJB-5-131 attenuated oxidative DNA damage, senescence and age-related pathology. Our findings indicate that nuclear genotoxic stress arises, at least in part, due to mitochondrial-derived ROS, and this spontaneous DNA damage is sufficient to drive increased levels of ROS, cellular senescence, and the consequent age-related physiological decline.
Keywords: Aging; Cellular senescence; Endogenous DNA damage; Free radicals; Genotoxic stress; Oxidative lesions; Reactive oxygen species.
Copyright © 2018 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Farr J.N., Xu M., Weivoda M.M., Monroe D.G., Fraser D.G., Onken J.L., Negley B.A., Sfeir J.G., Ogrodnik M.B., Hachfeld C.M., LeBrasseur N.K., Drake M.T., Pignolo R.J., Pirtskhalava T., Tchkonia T., Oursler M.J., Kirkland J.L., Khosla S. Targeting cellular senescence prevents age-related bone loss in mice. Nat. Med. 2017;23(9):1072–1079. - PMC - PubMed
-
- Jeon O.H., Kim C., Laberge R.M., Demaria M., Rathod S., Vasserot A.P., Chung J.W., Kim D.H., Poon Y., David N., Baker D.J., van Deursen J.M., Campisi J., Elisseeff J.H. Local clearance of senescent cells attenuates the development of post-traumatic osteoarthritis and creates a pro-regenerative environment. Nat. Med. 2017;23(6):775–781. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AG044376/AG/NIA NIH HHS/United States
- U19 AI068021/AI/NIAID NIH HHS/United States
- R01 CA076541/CA/NCI NIH HHS/United States
- P30 CA047904/CA/NCI NIH HHS/United States
- R29 CA076541/CA/NCI NIH HHS/United States
- K99 AG049126/AG/NIA NIH HHS/United States
- R01 CA101864/CA/NCI NIH HHS/United States
- R01 HL142248/HL/NHLBI NIH HHS/United States
- U19 AG056278/AG/NIA NIH HHS/United States
- P30 AG044271/AG/NIA NIH HHS/United States
- R00 AG049126/AG/NIA NIH HHS/United States
- P01 AG043376/AG/NIA NIH HHS/United States
- P30 AG024827/AG/NIA NIH HHS/United States
- P20 GM109098/GM/NIGMS NIH HHS/United States
- R00 AG036817/AG/NIA NIH HHS/United States
- R01 ES016114/ES/NIEHS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
