

The capacity of oocytes for DNA repair
- PMID: 29748894
- PMCID: PMC11105623
- DOI: 10.1007/s00018-018-2833-9
The capacity of oocytes for DNA repair
Abstract
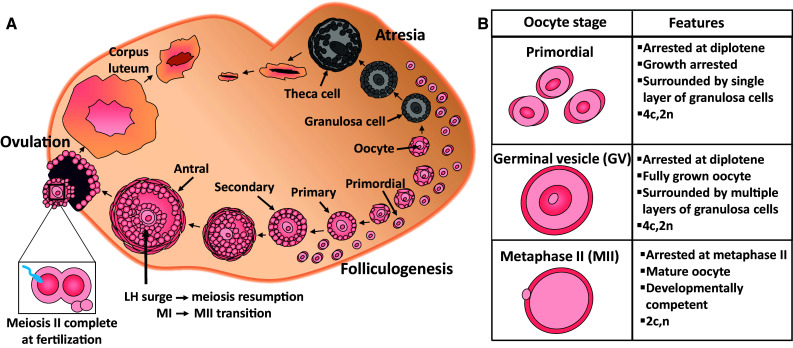
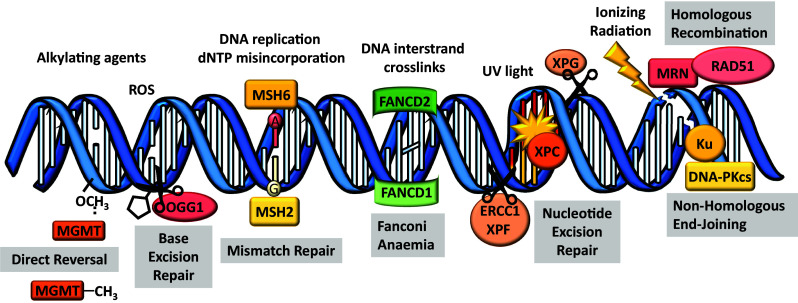
Female fertility and offspring health are critically dependent on the maintenance of an adequate supply of high-quality oocytes. Like somatic cells, oocytes are subject to a variety of different types of DNA damage arising from endogenous cellular processes and exposure to exogenous genotoxic stressors. While the repair of intentionally induced DNA double strand breaks in gametes during meiotic recombination is well characterised, less is known about the ability of oocytes to repair pathological DNA damage and the relative contribution of DNA repair to oocyte quality is not well defined. This review will discuss emerging data suggesting that oocytes are in fact capable of efficient DNA repair and that DNA repair may be an important mechanism for ensuring female fertility, as well as the transmission of high-quality genetic material to subsequent generations.
Keywords: Base excision repair; Detection and response; Folliculogenesis; Homologous recombination; Mismatch repair; Non-homologous end joining; Nucleotide excision repair; Ovary; Primordial follicles.
Figures
Similar articles
-
The importance of DNA repair for maintaining oocyte quality in response to anti-cancer treatments, environmental toxins and maternal ageing.Hum Reprod Update. 2018 Mar 1;24(2):119-134. doi: 10.1093/humupd/dmy002. Hum Reprod Update. 2018. PMID: 29377997
-
Xrcc5/KU80 is not required for the survival or activation of prophase-arrested oocytes in primordial follicles.Front Endocrinol (Lausanne). 2023 Oct 10;14:1268009. doi: 10.3389/fendo.2023.1268009. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 37900135 Free PMC article.
-
Oocytes can efficiently repair DNA double-strand breaks to restore genetic integrity and protect offspring health.Proc Natl Acad Sci U S A. 2020 May 26;117(21):11513-11522. doi: 10.1073/pnas.2001124117. Epub 2020 May 7. Proc Natl Acad Sci U S A. 2020. PMID: 32381741 Free PMC article.
-
BRCA-related ATM-mediated DNA double-strand break repair and ovarian aging.Hum Reprod Update. 2020 Jan 1;26(1):43-57. doi: 10.1093/humupd/dmz043. Hum Reprod Update. 2020. PMID: 31822904 Free PMC article. Review.
-
Advances in the study of DNA damage and repair in mammalian oocytes.Yi Chuan. 2023 May 20;45(5):379-394. doi: 10.16288/j.yczz.23-018. Yi Chuan. 2023. PMID: 37194586 Review.
Cited by
-
Decreased fatty acids induced granulosa cell apoptosis in patients with diminished ovarian reserve.J Assist Reprod Genet. 2022 May;39(5):1105-1114. doi: 10.1007/s10815-022-02462-8. Epub 2022 Mar 26. J Assist Reprod Genet. 2022. PMID: 35347502 Free PMC article.
-
Sperm DNA integrity does play a crucial role for embryo development after ICSI, notably when good-quality oocytes from young donors are used.Biol Res. 2022 Dec 26;55(1):41. doi: 10.1186/s40659-022-00409-y. Biol Res. 2022. PMID: 36572948 Free PMC article.
-
The role of cellular senescence in female reproductive aging and the potential for senotherapeutic interventions.Hum Reprod Update. 2022 Feb 28;28(2):172-189. doi: 10.1093/humupd/dmab038. Hum Reprod Update. 2022. PMID: 34918084 Free PMC article. Review.
-
DNA damage in aging, the stem cell perspective.Hum Genet. 2020 Mar;139(3):309-331. doi: 10.1007/s00439-019-02047-z. Epub 2019 Jul 19. Hum Genet. 2020. PMID: 31324975 Free PMC article. Review.
-
Selective utilization of non-homologous end-joining and homologous recombination for DNA repair during meiotic maturation in mouse oocytes.Cell Prolif. 2023 Apr;56(4):e13384. doi: 10.1111/cpr.13384. Epub 2022 Dec 23. Cell Prolif. 2023. PMID: 36564861 Free PMC article.
References
-
- Jackson SP, Bartek J (2009) The DNA-damage response in human biology and disease. Nature 461(7267):1071–1078. http://www.nature.com/nature/journal/v461/n7267/suppinfo/nature08467_S1.... - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
