

Mycobacteria employ two different mechanisms to cross the blood-brain barrier
- PMID: 29749044
- PMCID: PMC6175424
- DOI: 10.1111/cmi.12858
Mycobacteria employ two different mechanisms to cross the blood-brain barrier
Abstract
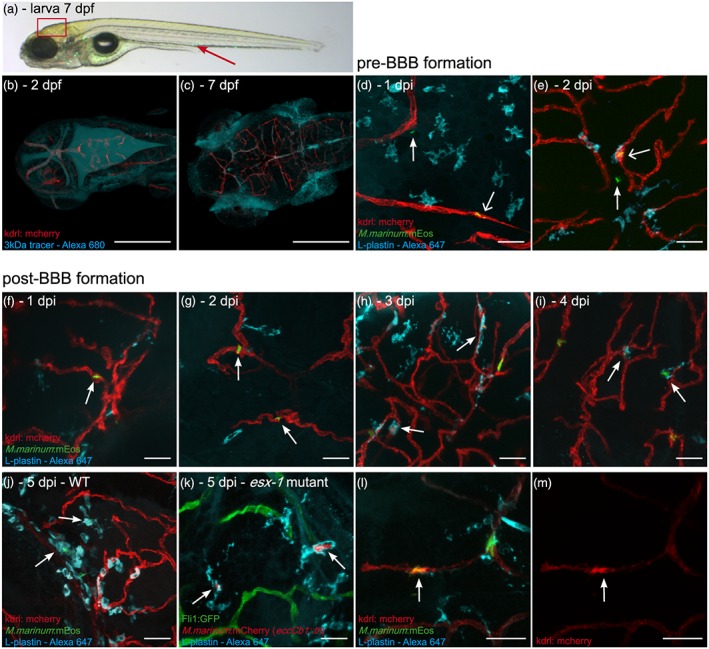
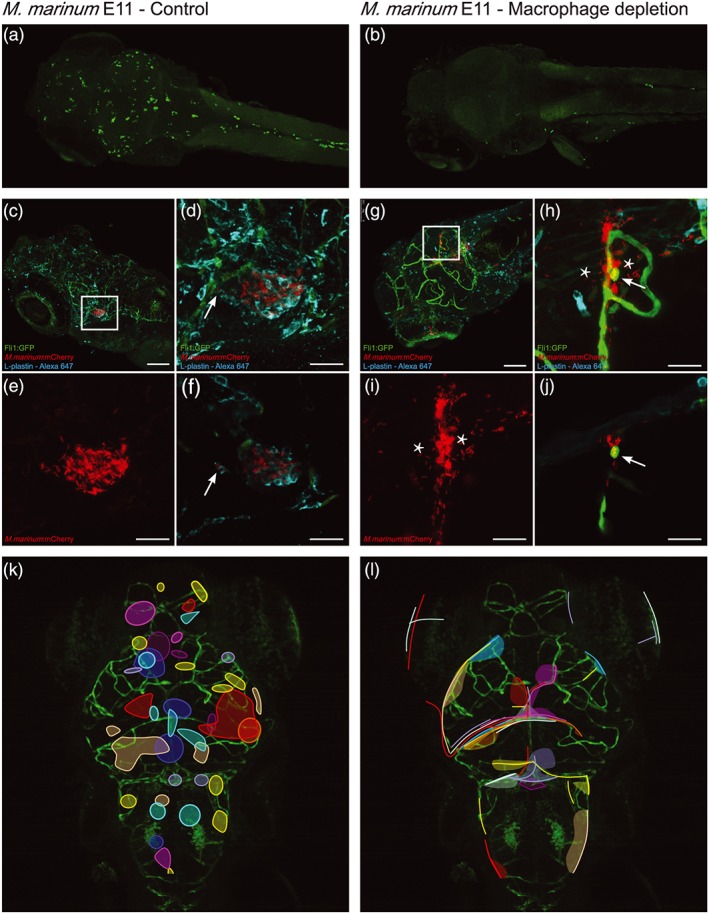
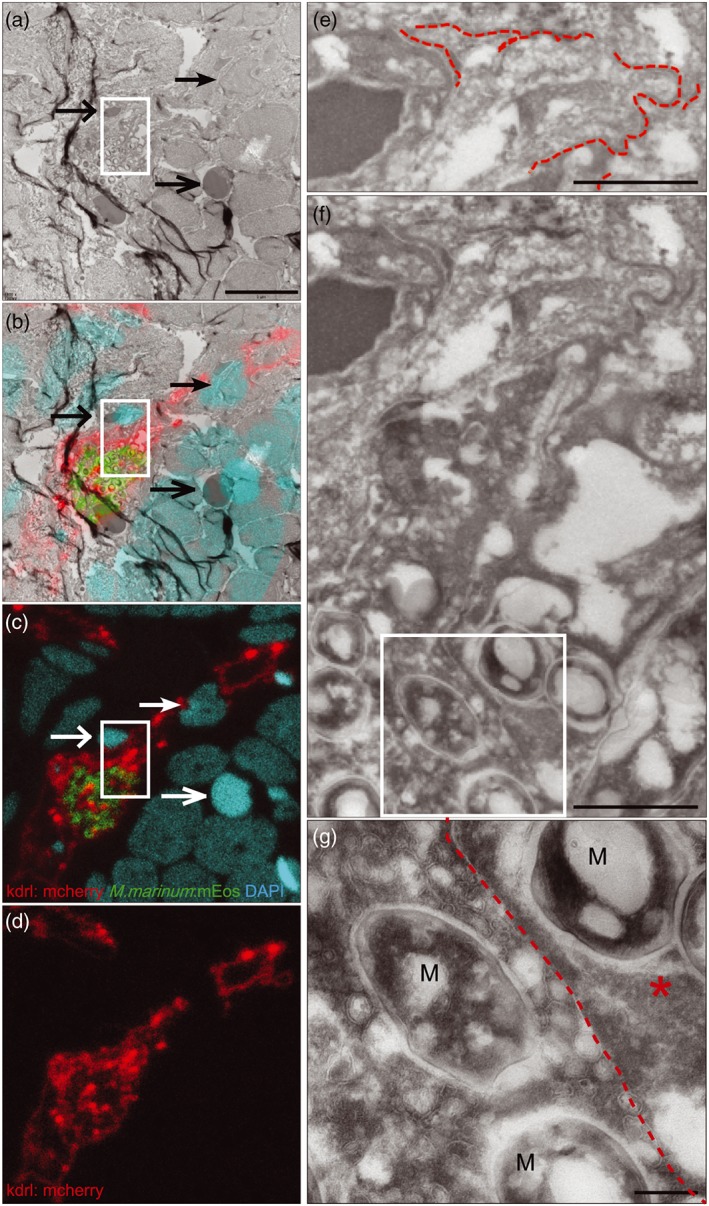
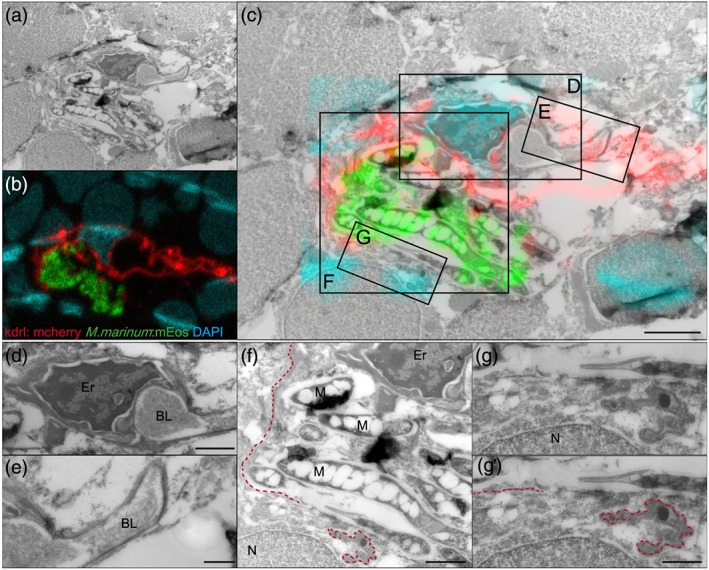
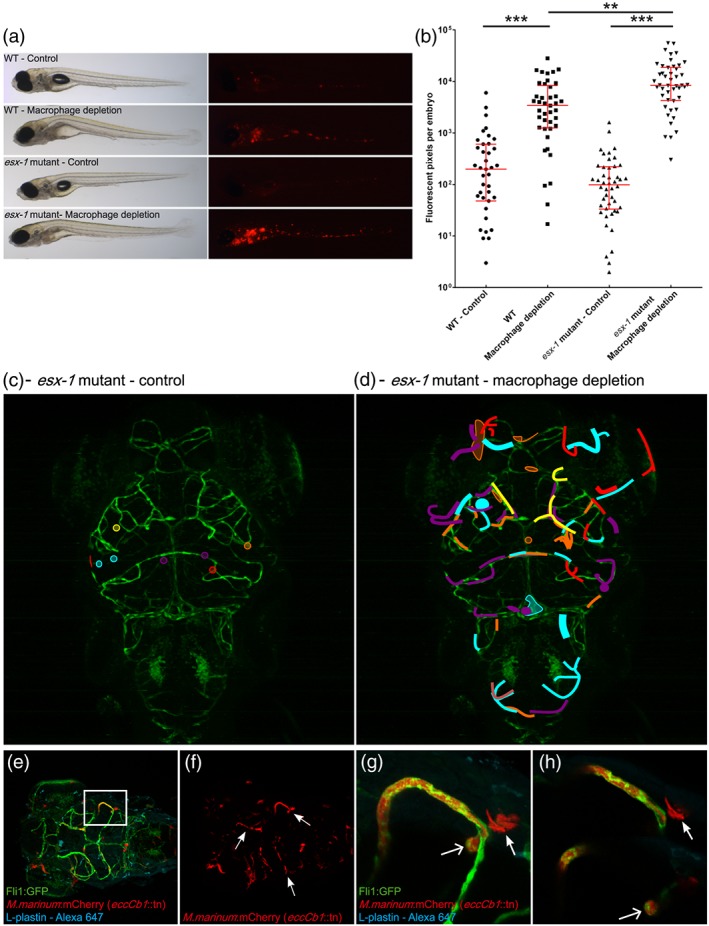
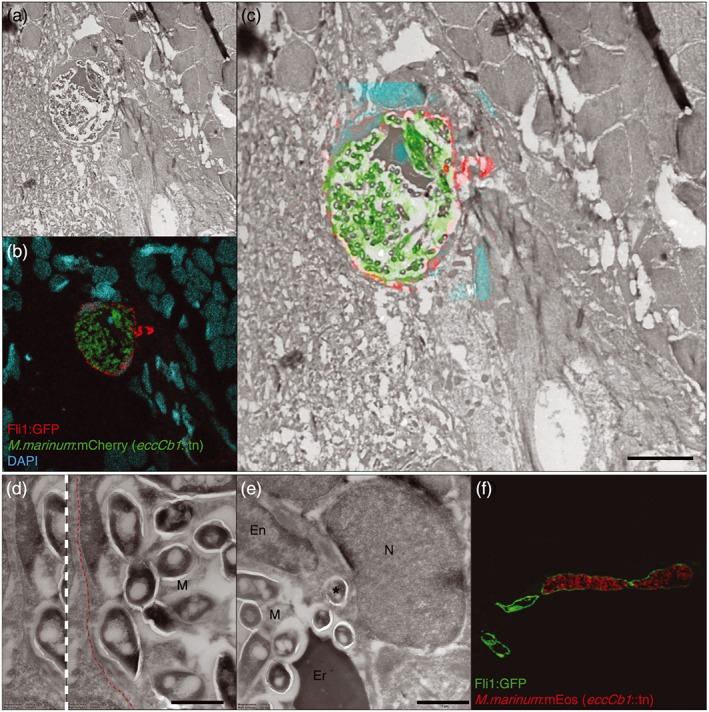
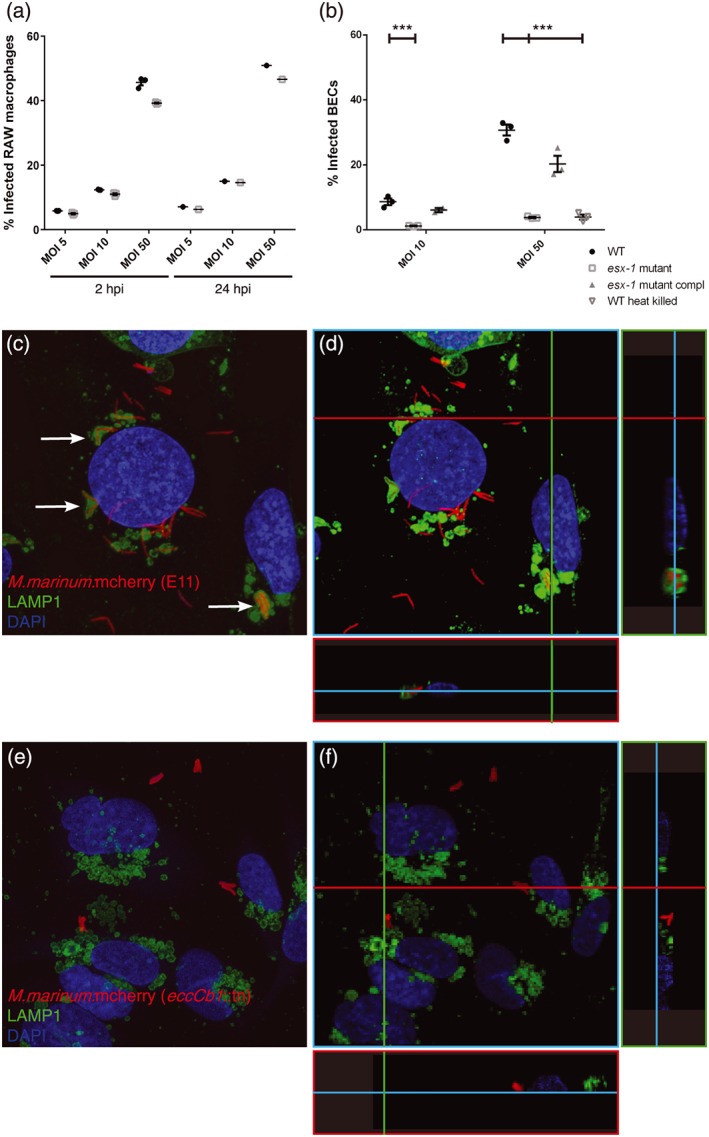
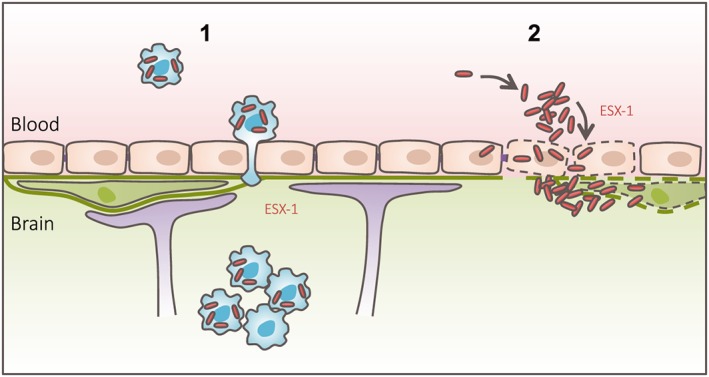
Central nervous system (CNS) infection by Mycobacterium tuberculosis is one of the most devastating complications of tuberculosis, in particular in early childhood. In order to induce CNS infection, M. tuberculosis needs to cross specialised barriers protecting the brain. How M. tuberculosis crosses the blood-brain barrier (BBB) and enters the CNS is not well understood. Here, we use transparent zebrafish larvae and the closely related pathogen Mycobacterium marinum to answer this question. We show that in the early stages of development, mycobacteria rapidly infect brain tissue, either as free mycobacteria or within circulating macrophages. After the formation of a functionally intact BBB, the infiltration of brain tissue by infected macrophages is delayed, but not blocked, suggesting that crossing the BBB via phagocytic cells is one of the mechanisms used by mycobacteria to invade the CNS. Interestingly, depletion of phagocytic cells did not prevent M. marinum from infecting the brain tissue, indicating that free mycobacteria can independently cause brain infection. Detailed analysis showed that mycobacteria are able to cause vasculitis by extracellular outgrowth in the smaller blood vessels and by infecting endothelial cells. Importantly, we could show that this second mechanism is an active process that depends on an intact ESX-1 secretion system, which extends the role of ESX-1 secretion beyond the macrophage infection cycle.
Keywords: ESX-1 secretion; Trojan horse mechanism; blood-brain barrier; tuberculosis; tuberculous meningitis; zebrafish.
© 2018 The Authors Cellular Microbiology Published by John Wiley & Sons Ltd.
Figures
References
-
- Abbott, N. J. , Rönnbäck, L. , & Hansson, E. (2006). Astrocyte‐endothelial interactions at the blood‐brain barrier. Nature Reviews. Neuroscience, 7(1), 41–53. Available at: http://www.ncbi.nlm.nih.gov/pubmed/16371949. Accessed May 22, 2013 - PubMed
-
- Abdallah, A. M. , Bestebroer, J. , Savage, N. D. , de Punder, K. , van Zon, M. , Wilson, L. , … Peters, P. J. (2011). Mycobacterial secretion systems ESX‐1 and ESX‐5 play distinct roles in host cell death and inflammasome activation. Journal of Immunology, 187(9), 4744–4753. Available at: http://www.ncbi.nlm.nih.gov/pubmed/21957139. Accessed December 31, 2013 - PubMed
-
- Be, N. a. , Bishai, W. R. , & Jain, S. K. (2012). Role of Mycobacterium tuberculosis pknD in the pathogenesis of central nervous system tuberculosis. BMC Microbiology, 12, 7 Available at: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3322341&tool=p... - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
