

Expression of AhDREB1, an AP2/ERF Transcription Factor Gene from Peanut, Is Affected by Histone Acetylation and Increases Abscisic Acid Sensitivity and Tolerance to Osmotic Stress in Arabidopsis
- PMID: 29751673
- PMCID: PMC5983730
- DOI: 10.3390/ijms19051441
Expression of AhDREB1, an AP2/ERF Transcription Factor Gene from Peanut, Is Affected by Histone Acetylation and Increases Abscisic Acid Sensitivity and Tolerance to Osmotic Stress in Arabidopsis
Abstract
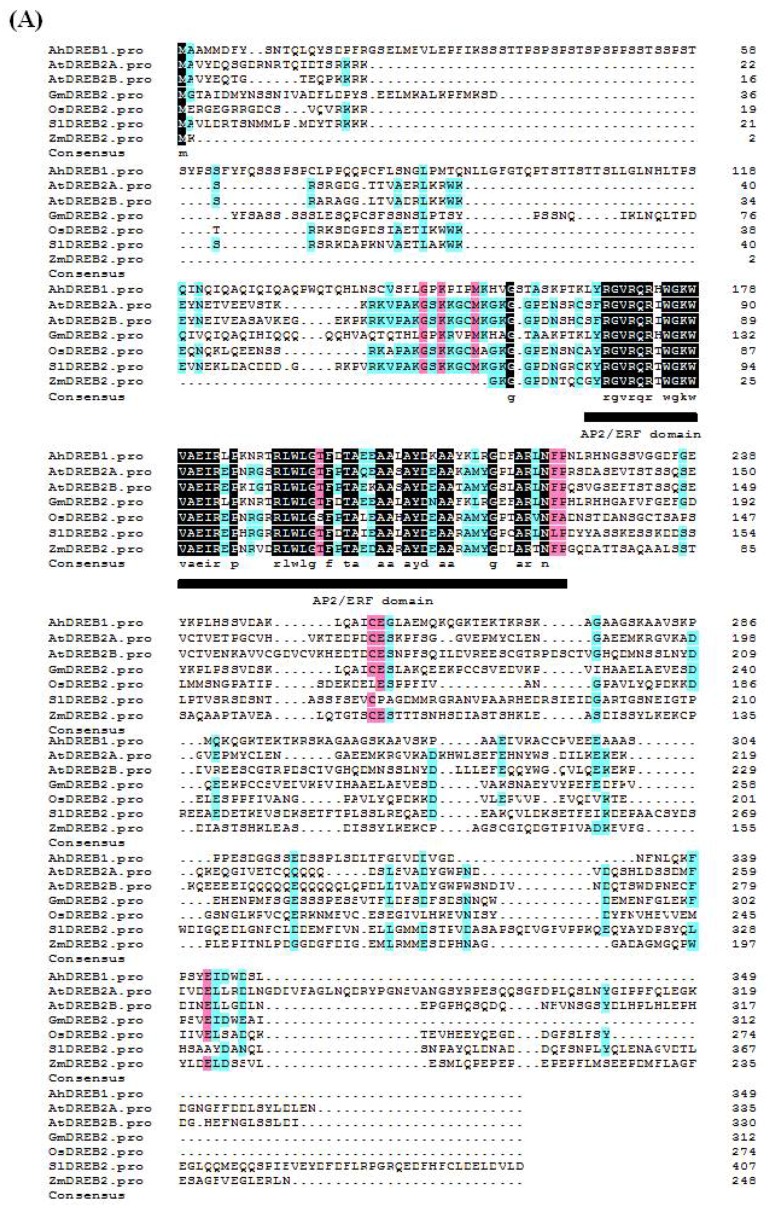
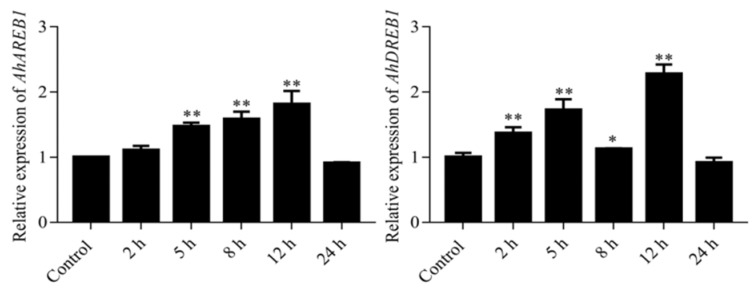
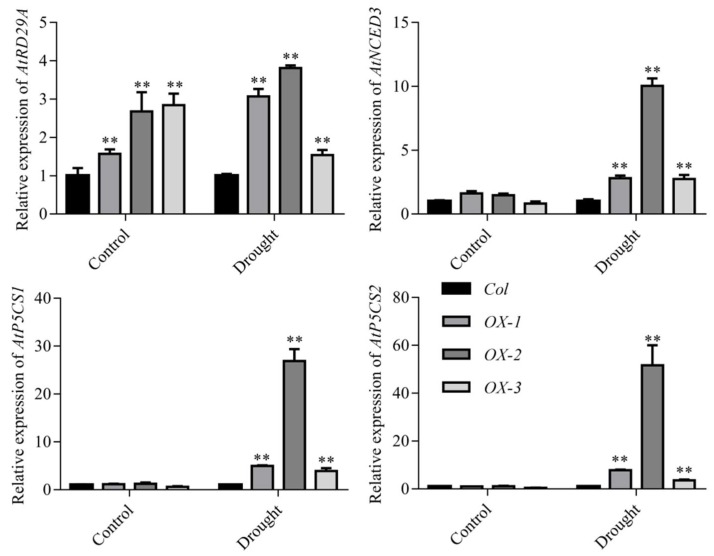
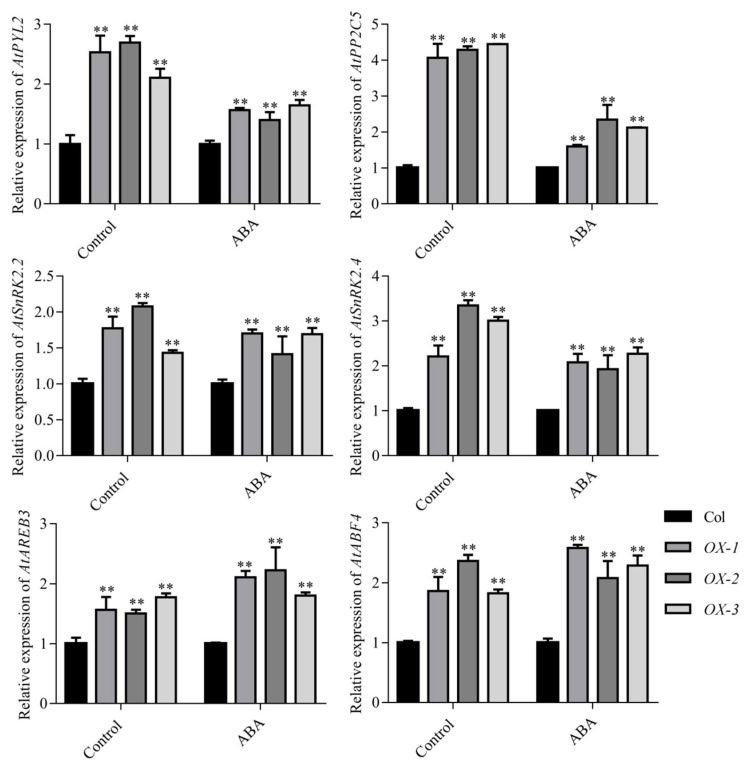
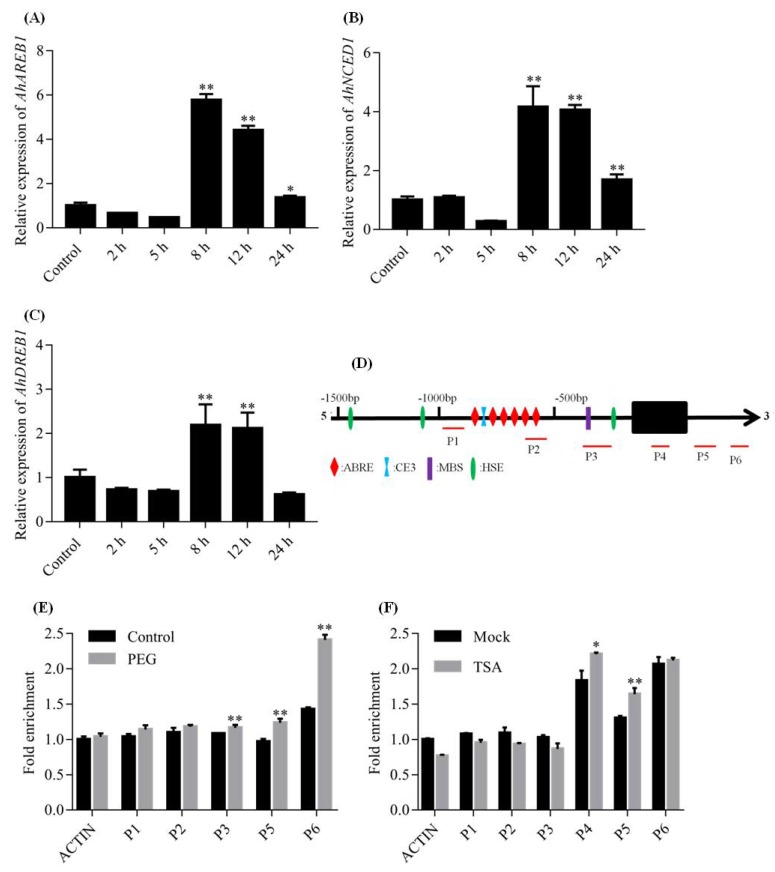
Drought stress negatively affects plant growth and development. An increasing number of reports have revealed the involvement of APETALA2/Ethylene Responsive Factor (AP2/ERF) transcription factors (TFs) in biotic and abiotic stress regulation in plants. However, research on these TFs in the peanut plant (Arachis hypogaea) has been limited. Here, we isolated a full-length coding sequence (CDS) of the AP2/ERF family gene AhDREB1 from the peanut plant and showed that its expression was induced by Polyethylene Glycol (PEG) 6000 and exogenous abscisic acid (ABA) treatment. When overexpressed in Arabidopsis, AhDREB1 increased both ABA levels and ABA sensitivity, affected the ABA signaling pathway and increased the expression of downstream drought stress-related genes RD29A, P5CS1, P5CS2 and NCED1. These results demonstrate that AhDREB1 can improve tolerance to drought via the ABA-dependent pathway in Arabidopsis. In the peanut plant, the specific histone deacetylases (HDACs) inhibitor trichostatin A (TSA) promotes AhDREB1 transcription and the enrichment level of H3ac was increased in regions of the AhDREB1 gene during TSA and PEG treatment. In summary, histone acetylation can affect the expression of AhDREB1 under osmotic stress conditions, thereby improving plant drought resistance.
Keywords: APETALA2/Ethylene Responsive Factor; AhDREB1; abscisic acid; drought stresses; histone acetylation; peanut.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





References
-
- Gonzalez-Villagra J., Rodrigues-Salvador A., Nunes-Nesi A., Cohen J.D., Reyes-Diaz M.M. Age-related mechanism and its relationship with secondary metabolism and abscisic acid in Aristotelia chilensis plants subjected to drought stress. Plant Physiol. Biochem. 2018;124:136–145. doi: 10.1016/j.plaphy.2018.01.010. - DOI - PubMed
-
- Bohnert H.J., Jensen R.G. Strategies for engineering water-stress tolerance in plants. Trends Biotechnol. 1996;14:89–97. doi: 10.1016/0167-7799(96)80929-2. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
