

Caspase-mediated cleavage of the centrosomal proteins during apoptosis
- PMID: 29752437
- PMCID: PMC5948218
- DOI: 10.1038/s41419-018-0632-8
Caspase-mediated cleavage of the centrosomal proteins during apoptosis
Abstract
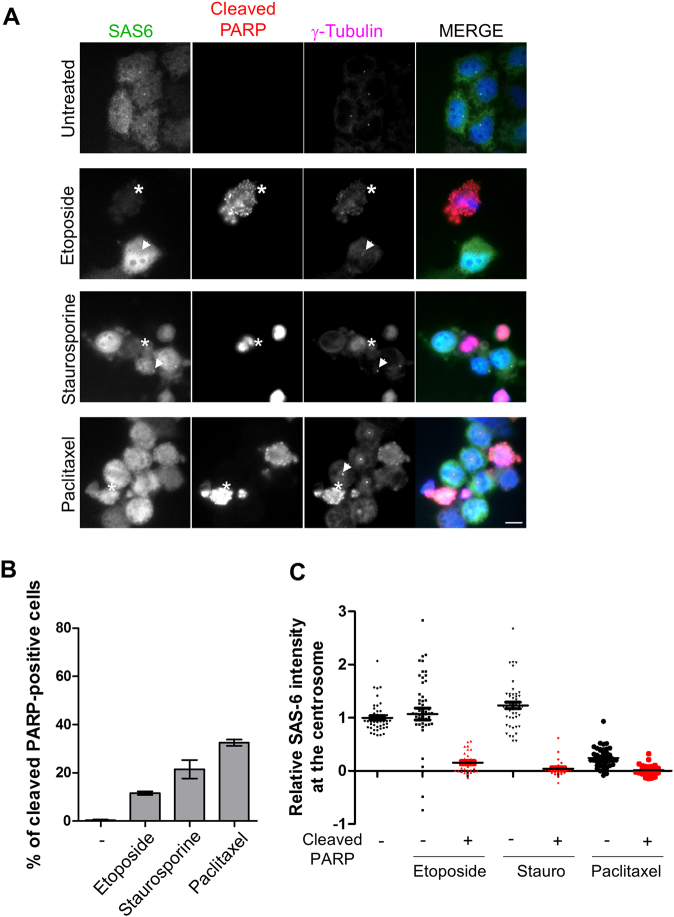
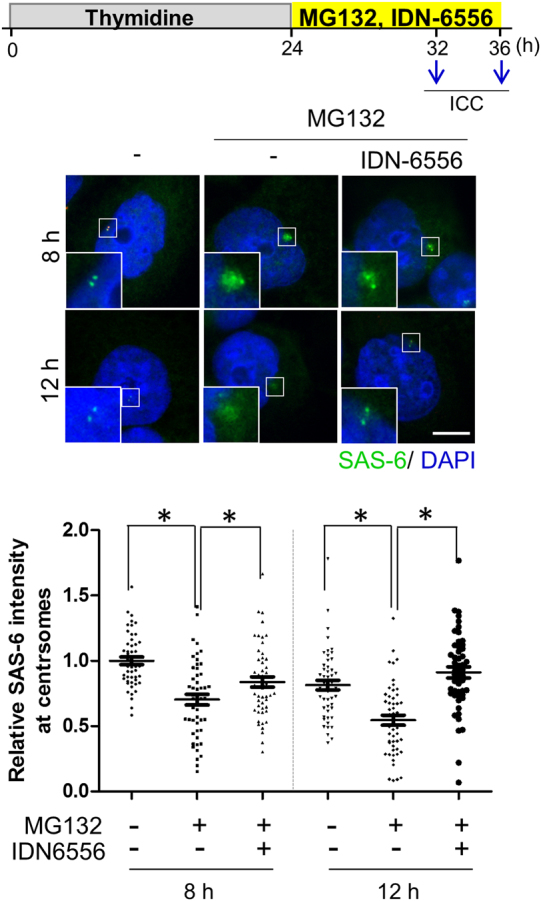
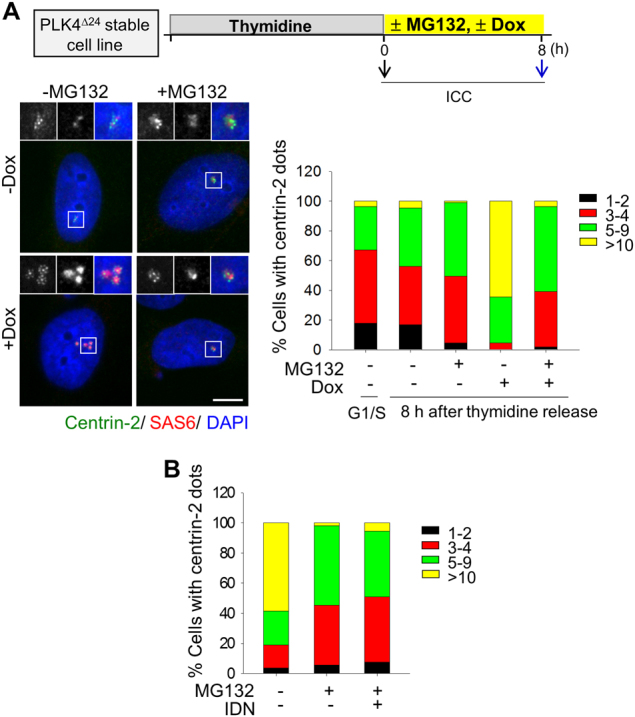
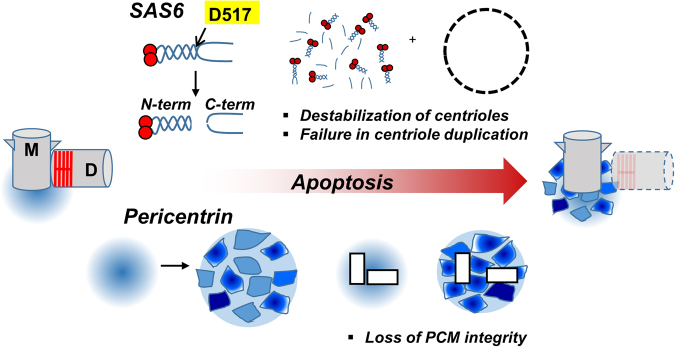
The centrosome is the major microtubule-organizing center and plays important roles in intracellular transport, cellular morphology, and motility. In mitotic cells, centrosomes function as spindle poles to pull a set of chromosomes into daughter cells. In quiescent cells, primary cilia are originated from the centrosomes. Given its involvement in various cellular processes, it is little surprising that the organelle would also participate in apoptotic events. However, it remains elusive how the centrosome changes in structure and organization during apoptosis. Apoptosis, a programmed cell death, is required for homeostatic tissue maintenance, embryonic development, stress responses, etc. Activation of caspases generates a cascade of apoptotic pathways, explaining much of what happens during apoptosis. Here, we report the proteolytic cleavage of selected centrosomal proteins in apoptotic cells. SAS-6, a cartwheel component of centrioles, was specifically cleaved at the border of the coiled-coil domain and the disordered C-terminus. Pericentrin, a scaffold of pericentriolar material, was also cleaved during apoptosis. These cleavages were efficiently blocked by the caspase inhibitors. We propose that the caspase-dependent proteolysis of the centrosomal proteins may destabilize the configuration of a centrosome. Loss of centrosomes may be required for the formation of apoptotic microtubule networks, which are essential for apoptotic fragmentation. This work demonstrates the first centrosomal targets by caspases during apoptosis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
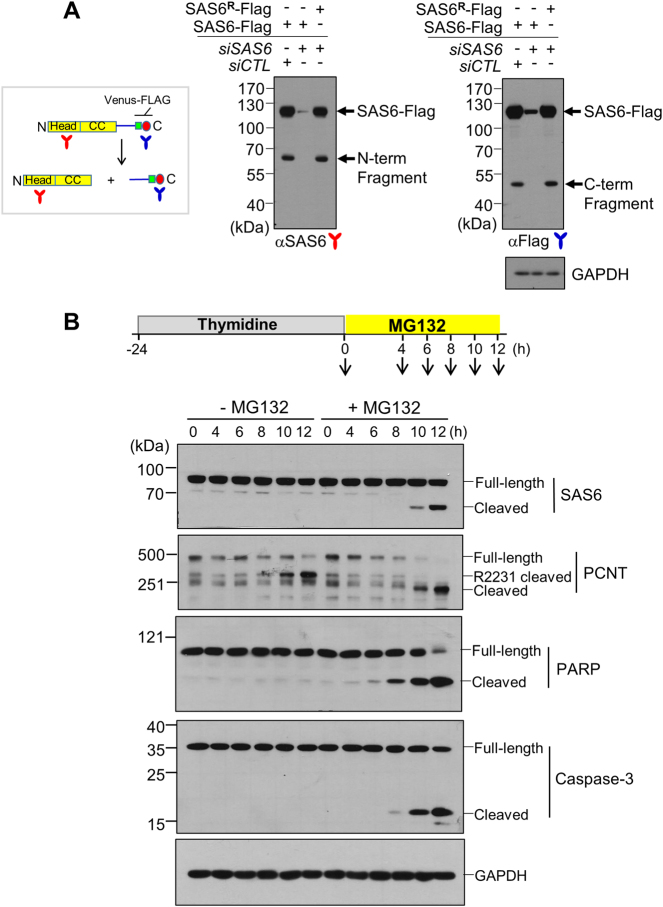
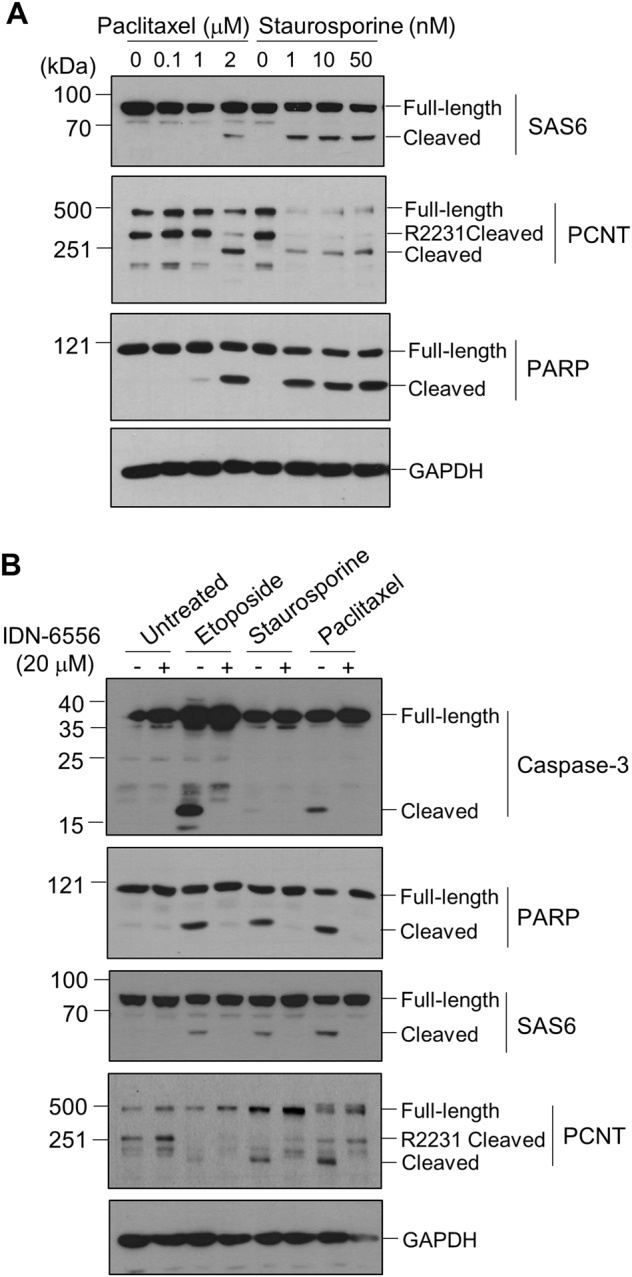
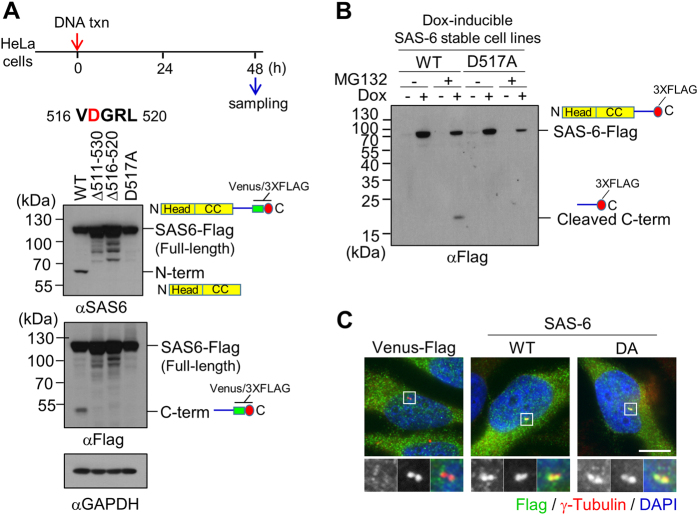
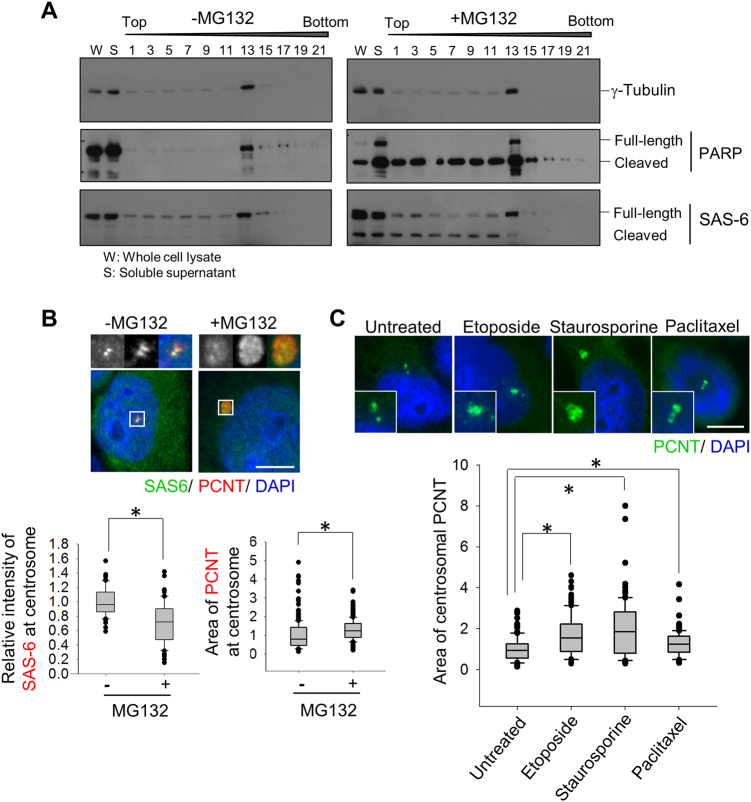
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
