

Membrane Potential Distinctly Modulates Mobility and Signaling of IL-2 and IL-15 Receptors in T Cells
- PMID: 29754714
- PMCID: PMC6129476
- DOI: 10.1016/j.bpj.2018.04.038
Membrane Potential Distinctly Modulates Mobility and Signaling of IL-2 and IL-15 Receptors in T Cells
Abstract
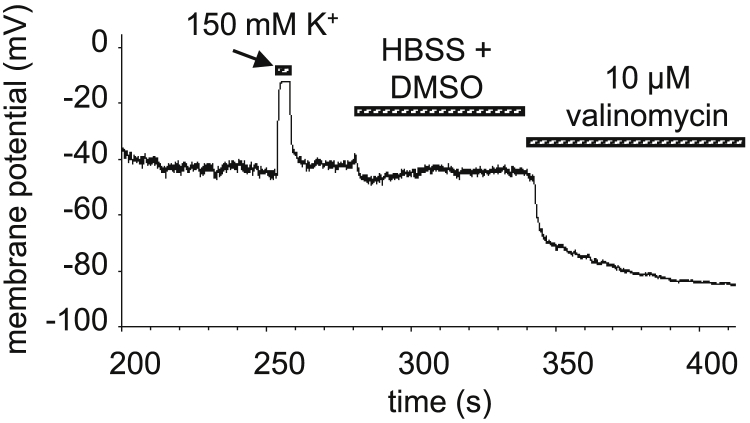
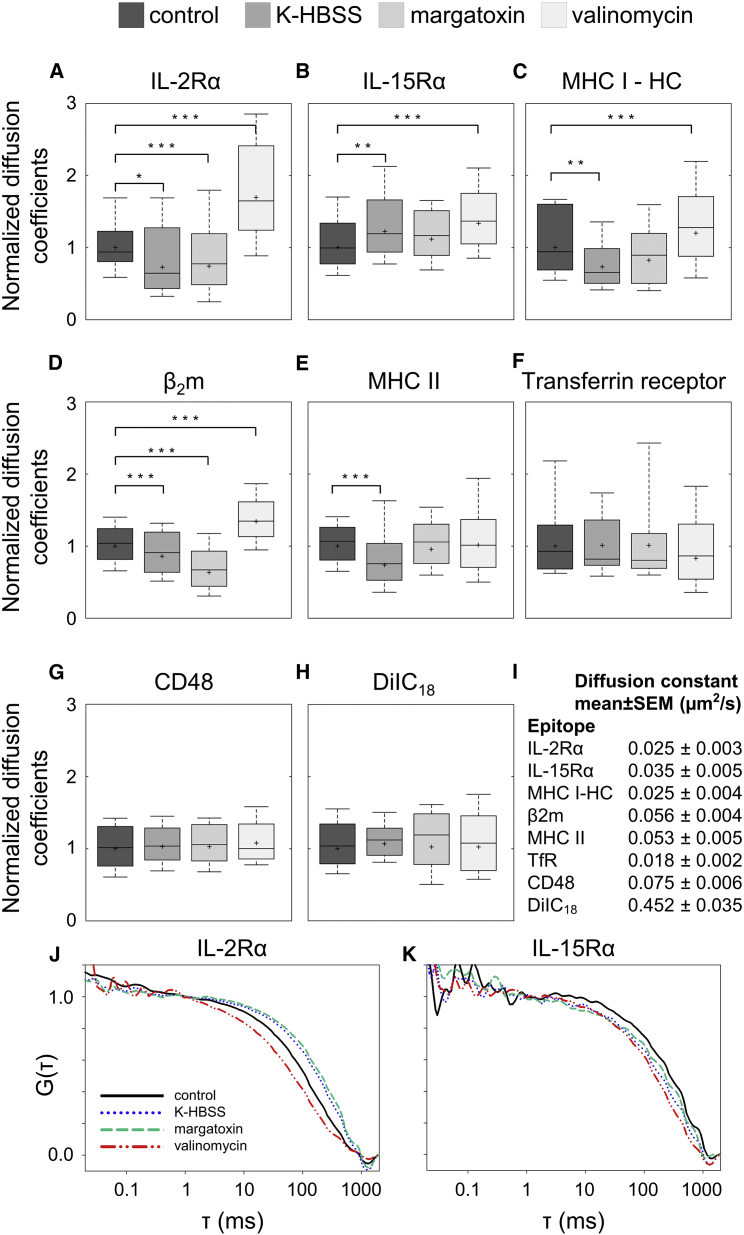
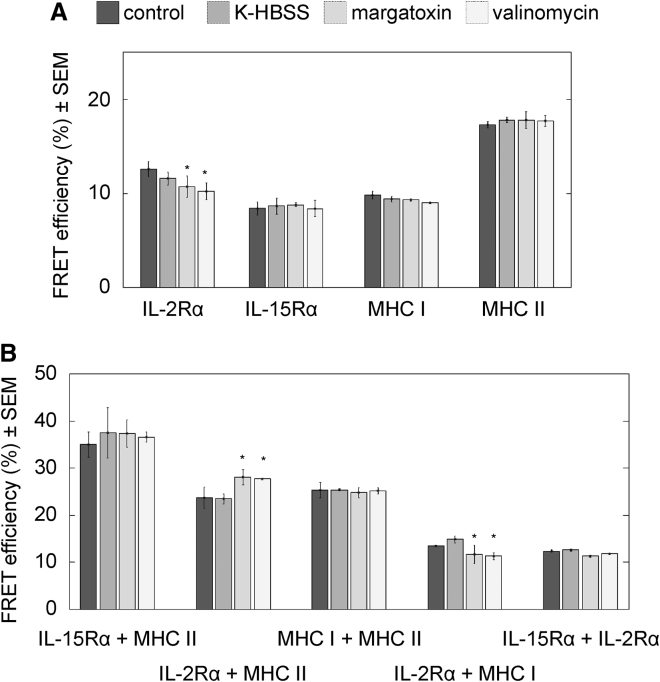
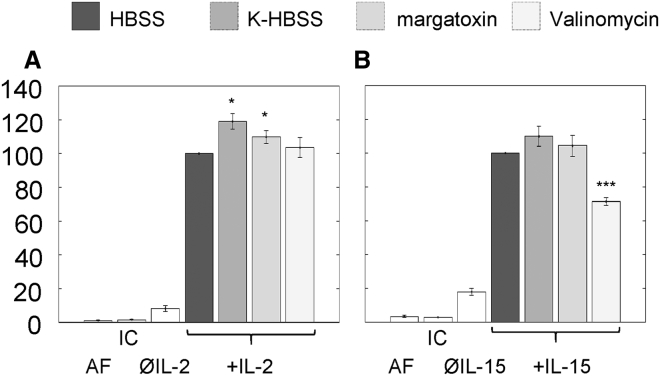
The high electric field across the plasma membrane might influence the conformation and behavior of transmembrane proteins that have uneven charge distributions in or near their transmembrane regions. Membrane depolarization of T cells occurs in the tumor microenvironment and in inflamed tissues because of K+ release from necrotic cells and hypoxia affecting the expression of K+ channels. However, little attention has been given to the effect of membrane potential (MP) changes on membrane receptor function. Therefore, we studied the influence of membrane de- and hyperpolarization on the biophysical properties and signaling of interleukin-2 (IL-2) and interleukin-15 (IL-15) receptors, which play important roles in T cell function. We investigated the mobility, clustering, and signaling of these receptors and major histocompatibility complex (MHC) I/II glycoproteins forming coclusters in lipid rafts of T cells. Depolarization by high K+ buffer or K+ channel blockers resulted in a decrease in the mobility of IL-2Rα and MHC glycoproteins, as shown by fluorescence correlation spectroscopy, whereas hyperpolarization by the K+ ionophore valinomycin increased their mobility. Contrary to this, the mobility of IL-15Rα decreased upon both de- and hyperpolarization. These changes in protein mobility are not due to an alteration of membrane fluidity, as evidenced by fluorescence anisotropy measurements. Förster resonance energy transfer measurements showed that most homo- or heteroassociations of IL-2R, IL-15R, and MHC I did not change considerably, either. MP changes modulated signaling by the two cytokines in distinct ways: depolarization caused a significant increase in the IL-2-induced phosphorylation of signal transducer and activator of transcription 5, whereas hyperpolarization evoked a decrease only in the IL-15-induced signal. Our data imply that the MP may be an important modulator of interleukin receptor signaling and dynamics. Enhanced IL-2 signaling in depolarized Treg cells highly expressing IL-2R may contribute to suppression of antitumor immune surveillance.
Copyright © 2018 Biophysical Society. All rights reserved.
Figures
References
-
- Fehniger T.A., Cooper M.A., Caligiuri M.A. Interleukin-2 and interleukin-15: immunotherapy for cancer. Cytokine Growth Factor Rev. 2002;13:169–183. - PubMed
-
- Waldmann T.A. The biology of interleukin-2 and interleukin-15: implications for cancer therapy and vaccine design. Nat. Rev. Immunol. 2006;6:595–601. - PubMed
-
- Waldmann T.A., Dubois S., Tagaya Y. Contrasting roles of IL-2 and IL-15 in the life and death of lymphocytes: implications for immunotherapy. Immunity. 2001;14:105–110. - PubMed
-
- Germain R.N. MHC-dependent antigen processing and peptide presentation: providing ligands for T lymphocyte activation. Cell. 1994;76:287–299. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
